Растения, входящие в отдел хвощевидных (Equisetophyta ), отличаются от других споровых побегами, составленными отдельными члениками (междоузлиями) и узлами с мутовчато расположенными листьями. Характерной чертой хвощевидных является также наличие у них спорангиофоров – особых структур, несущих спорангии. Мутовки спорангиофоров образуют на стебле спороносные зоны, чередующиеся с обычными вегетативными листьями, или сидят на концах осей, образуя стробилы.
Ископаемые остатки хвощевидных известны из девонских отложений. К концу этого периода хвощевидные уже прошли длительный путь эволюции, а расцвет этой группы растений пришелся на каменноугольное время. В палеозое хвощевидные отличались исключительным разнообразием: среди них встречались травы, кустарники, лианы и деревья, достигавшие 20 м высоты. Вместе с гигантскими плауновидными эти формы образовывали каменноугольные леса.
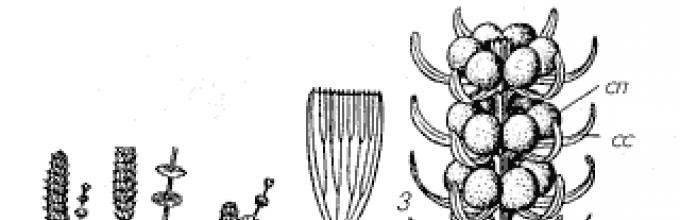
В то время, помимо представителей класса хвощевых (Equisetopsida ), на Земле произрастали и хвощевидные растения другого класса – клинолистовых (Sphenophyllopsida )1 . Клинолисты (род Sphenophyllum ) образовывали густые заросли в лесах каменноугольного периода (рис. 1). Побеги некоторых растений этой группы достигали в длину нескольких метров при очень небольшом диаметре (0,1–0,5 см) и были либо полегающими, либо лазающими, о чем свидетельствуют прицепки, обнаруженные на стеблях одного из видов. К узлам молодых участков стебля прикреплялись мутовки из 6–9 клиновидных листьев.

1,2 – клинолист клинолистный (Sphenophyllum cuneifolium) (1 – общий вид части растения, 2 – листья разных узлов стебля); 3–5 – стробилы представителей порядка клинолистовых
(3 – сфенофиллостахис водяной (Sphenophyllostachys aquensis ); 4–5 – сфенофиллостахис Досона (S. dawsonii ), (4 – продольный срез части стробила, 5 – поперечный срез стробила в области междоузлия; сп – спорангий, сс – стерильный сегмент спорангиофора, вс – веточка спороносного сегмента спорангиофора)
Древние представители класса хвощевых – каламиты (Calamites ) – в каменноугольном периоде в значительной мере составляли древесный ярус заболоченных тропических лесов. По внешнему виду каламиты напоминали современные хвощи, только гигантских размеров (рис. 2). Стволы каламитов достигали 10–20 м высоты и 0,5 м в диаметре за счет способности к вторичному утолщению (слой вторичной древесины в их стеблях достигал 12 см).
1–3 – каламиты (Calamites) с разными типами ветвления; 4 – листья каламитов; 5 – поперечный срез стробила каламита; 6 – часть стробила, вид сбоку: слева – с поверхности, справа – в продольном разрезе (б – брактея, пп – проводящий пучок, сф – спорангиофор, сп – спорангий)
Листья, основные фотосинтезирующие органы каламитов, длиной от 2 до 7 см, были расположены в узлах стебля мутовками по 3–70 штук. Стробилы, длиной около 12 и шириной около 4 см, состояли из чередующихся мутовок стерильных листьев и спорангиофоров. Возможно, все каламиты были разноспоровыми растениями, т.е. формировали мужские и женские споры, а на образовывавшихся из них гаметофитах (заростках), соответственно, развивались либо только мужские, либо только женские половые клетки. У одного из каламитов (каламокарпон замечательный – Calamocarpon insignis ) найден мегастробил, в спорангии которого развивалась всего одна мегаспора. Она прорастала прямо внутри спорангия, образуя женский гаметофит. Такая крайняя степень редукции числа мегаспор в спорангии отмечена и у древовидных плауновидных палеозоя 2 .
В отличие от большинства древовидных плауновидных и класса клиноситовых каламиты пережили каменноугольный период и были обильно представлены и в пермское время. Но к началу мезозойской эры каламитовые, достигшие высокой степени специализации, вымерли под воздействием резкого изменения климата. Их менее специализированные сородичи из класса хвощовых на некоторое время пережили климатические перемены и в первой половине мезозоя в некоторых областях Земли даже образовали почти чистые сообщества по берегам водоемов. Однако и они постепенно почти все вымерли, так что в наше время весь отдел хвощевидных представлен одним родом хвощ (Equisetum ), относящимся к классу хвощовых и включающим около 20 видов.
Латинское название Equisetum впервые употребил Плиний Старший, имея в виду, по-видимому, сходство ветвистых побегов хвоща с хвостом лошади (от лат. equius – лошадь и saeta, seta – щетина, жесткие волосы). В русском названии «хвощ» отражено сходство взрослого растения с хвостом или с пучком волос. В некоторых местностях России хвощи с мутовчато расположенными побегами называют «сосенка», «елка», а по наличию у некоторых видов бурых спороносных побегов, похожих на пест, – «пестиками» или «толкачиками».

1 – участки горизонтального и вертикального корневища с клубнями у хвоща полевого; 2 – то же у хвоща болотного; 3 – схема поперечного среза клубня хвоща полевого; 4 – схема поперечного среза корня хвоща полевого (вп – воздушная полость, пп – проводящий пучок, пт – покровная ткань)
Все современные хвощи – многолетние корневищные травы с мутовками бурых редуцированных листьев, утративших хлорофилл (не следует путать с листьями мутовки зеленых веточек в узлах стеблей). Корневища у всех хвощей глубоко залегающие, сильно разветвленные, в их узлах часто закладываются почки. Почки на корневищах нередко образуют короткие утолщенные междоузлия (называемые клубнями), складывающиеся в короткие цепочки, богатые крахмалом. От узлов корневищ отходят многочисленные придаточные корни двух типов: растущие вниз, часто утолщенные, и направленные вверх или в стороны (рис. 3). Стебли прямостоячие или восходящие, простые или мутовчато-ветвистые, цилиндрические или с продольными ребрами, состоят из узлов и полых междоузлий. В узлах побегов находятся листовые влагалища, по верхнему краю которых расположены зубцевидные листовые пластины (рис. 4, В , Г ; 6). Листья хвощей срастаются основаниями в кольцо, сидящее на узле и прикрывающее растущую зону междоузлий, богатую сахаром. Верхушки листьев имеют форму узких зубцов. В почке эти зубцы сращены и образуют колпачок, который при росте стебля разрывается на верхушке. В дальнейшем при росте стебля в длину и толщину, листовые пластинки отходят друг от друга, но разрыв происходит иногда не по всем линиям срастания, поэтому число зубцов по краю влагалища может не соответствовать действительному числу листьев. Форма влагалища, цвет, форма и время жизни зубцов различны у разных видов, и эти признаки используются при определении хвощей.

А – вегетативный побег; Б – спороносный побег; В, Г – листовые влагалища; Д – спорангиофор со спорангиями; Е–З – споры (1 – корневище с клубнями, 2 – стробил; 3 – щиток; 4 – спорангий; 5 – ножка; 6 – наружная оболочка споры, 7 – элатеры)
В основании листовых влагалищ хвощей закладываются почки, которые позже прорывают влагалища и образуют боковые побеги. Для возобновления растения особенно важны почки на тех узлах стеблей, которые находятся под землей, вблизи ее поверхности. Рядом с почкой закладывается корень.
По характеру надземных побегов хвощи можно разбить на две группы. У одних видов все надземные побеги однотипны. Они очень жесткие, обычно вечнозеленые (т.е. живут более одного сезона и в умеренной зоне перезимовывают) и развивают верхушечные стробилы.
У видов второй группы побеги бывают двух типов: одни – спороносные – буроватые или зеленые, а другие – вегетативные, зеленые. Побеги обоих типов появляются весной над поверхностью почвы, они нежнее по своей консистенции, чем побеги первой группы, и к зиме отмирают. Развитие вегетативных и спороносных побегов у видов второй группы идет по-разному. У хвоща полевого спороносные побеги появляются первыми ранней весной (рис. 4, В ). Они буровато-розовые, неветвистые, обычно толще быстро зеленеющих вегетативных побегов и после спороношения большей частью отмирают. У хвощей лесного и лугового спороносные побеги с хорошо развитыми стробилами появляются ранней весной одновременно с вегетативными. И те и другие вначале бледно-розовые, а затем зеленеют. После спороношения и отсыхания стробила спороносные побеги функционируют подобно вегетативным. Наконец, у таких видов, как хвощ болотный, хвощ приречный, хвощ боготский (E. bogotense – он обитает в тропиках Центральной и Южной Америки), спороносные побеги в момент их появления над поверхностью земли с трудом можно отличить от вегетативных – у них такая же зеленая окраска и сходные размеры.

Рис. 5. Формы листовых мутовок (а) и поперечные срезы побегов (б) хвощей:
1 – хвощ полевой; 2 – хвощ луговой; 3 – хвощ лесной; 4 – хвощ болотный; 5 – хвощ зимующий (а – листовое кольцо; б – поперечный разрез стебля)
Жесткость побегов хвощей объясняется тем, что в состав оболочки клеток эпидермы и, в меньшей степени, других тканей побега наряду с целлюлозой входит кремнезем. Частицы кремнезема располагаются в клеточной оболочке между тяжами целлюлозы, а на поверхности клеток эпидермы образуют сплошной и гомогенный, очень прочный слой с мелкими бугорками, приуроченными к выростам оболочки (рис. 5). Слой из кремнезема покрыт снаружи тонкой кутикулой с восковым налетом и играет не только механическую, но и защитную роль, так как благодаря ему хвощи практически не подвергаются нападению различных растительноядных моллюсков и насекомых.
Стробилы или спороносные колоски хвощей состоят из спорангиофоров, размещенных на общей оси. Они имеют вид шестиугольных щитков с ножками (рис. 4, Д ). На внутренней стороне такого щитка размещены 4–6 спорангиев. Когда находящиеся в них споры созревают, ось колоска немного удлиняется, и щитки спорангиофоров отходят друг от друга. Стенки созревших спорангиев лопаются, открывается продольная щель, и споры высыпаются. Лентовидные придатки (элатеры), образующиеся из наружной оболочки споры, способствуют сцеплению распространяющихся спор друг с другом (рис. 4, Ж , З ).
Споры могут прорастать только на затененной и влажной почве, где из них вырастает пластинчатый или ветвистый заросток. Заростки (гаметофиты) современных хвощей – очень маленькие, размером в несколько миллиметров, недолговечные зеленые растеньица.
Современные хвощи в отличие от вымерших каламитов – равноспоровые растения, на их заростках образуются органы (архегонии и антеридии), продуцирующие как женские (яйцеклетки), так и мужские (многожгутиковые сперматозоиды) половые клетки.
Сперматозоиды хвощей подвижны, поэтому оплодотворение происходит в воде – во время дождя или выпадения росы. Из зиготы развивается зародыш, а из него – без периода покоя – проросток, который через несколько месяцев превращается во взрослую особь нового бесполого поколения – спорофит (рис. 6).

Рис. 6. Чередование поколений в жизненном цикле хвощей (Equisetum arvense):
а – гаметофит (заросток); б – спорофит; в – спорангофор; г – спорангии; д – щиток спорангиофора; е – элатеры; ж – клубеньки
В природе хвощи обычно встречаются в виде клонов (групп растений, возникших путем вегетативного размножения), которые нередко занимают участки площадью в несколько десятков и даже сотен квадратных метров.
Хвощи отличаются крайним полиморфизмом. Например, у хвоща полевого, недавно поселившегося на хорошо освещенном участке с нарушенным растительным покровом, стебли обычно низкие, распростертые. Но позже, когда разовьется окружающая растительность, от более зрелых корневищ того же клона могут отходить мощные прямостоячие стебли с ветвистыми побегами. При приближении к северным границам ареала этого вида спороносные побеги, буроватые у растений умеренной зоны, сменяются зеленеющими, а потом – в тундре – и ветвистыми зелеными. Очень изменчивы бывают побеги и у других видов. Эти модификации, возникающие под влиянием многообразных факторов внешней среды (часто даже в пределах одного клона), наряду с многочисленными гибридами и наследуемыми уродствами, являются одной из причин разногласий между ботаниками в оценке объема рода.
Хвощи довольно широко распространены на Земле, особенно в Северном полушарии. Но в Австралии, Новой Зеландии и Тропической Африке аборигенных видов хвощей нет.
Встречаются хвощи в различных растительных сообществах, но обязательно с достаточным или избыточным увлажнением. По берегам водоемов, в сырых лесах, в некоторых типах низинных болот хвощи нередко доминируют в травянистом покрове.
Хвощи часто захватывают территории с нарушенным растительным покровом, образуя при этом чистые или почти чистые заросли в тех местах, где другие растения не могут жить, например, из-за обилия воды. Раз поселившись на какой-нибудь территории, хвощи, благодаря наличию глубоко залегающих корневищ, масса которых превышает массу надземных частей растения в несколько раз, успешно противостоят таким неблагоприятным воздействиям внешней среды, как засухи, лесные пожары и т.п., и успешно конкурируют с другими растениями, зачастую становясь широко распространенными и трудно искореняемыми сорняками пастбищ и полей.
Хвощи, растущие в условиях избыточного увлажнения, обладают признаками гигрофитов – у них слабо развита водопроводящая система, но хорошо представлена воздухоносная ткань. Одновременно эти растения могут нести и признаки ксерофитов – погруженные под поверхность эпидермы замыкающие клетки устьиц, закрытые устьица на старых участках стебля, сильно развитая механическая ткань, фотосинтезирующие стебли, редуцированные листья. Соотношение тех или иных признаков строения варьирует у разных видов.
В умеренной зоне России чаще всего можно встретить следующие виды хвощей.

Рис. 7. Хвощ луговой (Equisetum pratense Ehrh.)авт. рис. Шикина А.В.
Хвощ луговой (E.pratense, рис. 7) – многолетнее растение с удлиненным подземным черным корневищем без клубней. Спороносные побеги бледно-зеленые, простые, с воронковидными крупными влагалищами и свободными светло-коричневыми зубцами, с перепончатой каймой. Колосок длиной 0,8–2 см, цилиндрический. Спороношение в июне-июле. После спороношения, в начале лета, из спороносных побегов развиваются простые зеленые ветви, и они становятся похожими на вегетативные. Вегетативные побеги хвоща лугового имеют высоту 10–30 см и диаметр около 2 мм, они прямостоячие, также бледно-зеленые, внутри с большой центральной полостью и многими мелкими периферическими (рис. 5), с 8–14 шероховатыми ребрами, с простыми неветвящимися боковыми трехгранными веточками длиной 5–15 см, расположенными горизонтально или дугообразно книзу. Зубцы листовых мутовок широколанцетные, на ветвях яйцевидные, острые, в количестве 10–15 шт.
Продолжение следует
Побеги хвощевидных имеют членистое строение, состоят из узлов и междоузлий. Листья собраны в мутовки.
К хвощевидным принадлежат как травянистые растения (из ныне живущих и вымерших) со стеблем от нескольких сантиметров до нескольких метров, так и древовидные (только вымершие), достигавшие 15 м и в диаметре более 0,5 м.
Проводящая система стебля хвощевидных - актиностела или артростела. Большинство хвощевидных - равноспоровые растения, и лишь некоторые ископаемые формы были разноспоровыми.
Отдел хвощевидные объединяет два класса: клинолистовые (Sphenophyllopsida) и хвощевые (Equisetopsida).
Ранее относимый класс гиениевые с преджставителями протогиения (Protohyenia), гиения (Hyenia, рис. 25) и каламофитон (Colamophyton) в настоящее время рассматриваются палеоботаниками как древнейшие кладоксиловые папоротники. У каламофитона описанные ранее узлы членистого побега оказались просто поперечными трещинами в породе. Анатомическое строение гиении пока неизвестно, а спороносные органы обоих видов редко отличаются от спорангиофоров девонских хвощевидных (Мейен: Еленевский и др. 2000).
Класс хвощовые (EQUISETOPSIDA)
Класс хвощовые содержит п о р я д о к хвощовые (Equisetales) , с е - м е й с т в а каламитовые (Calamitaceae) и хвощовые (Equisetaceae).
Вымершие представители объединены в семейство каламитовые. Виды этого семейства были широко распространены в карбоне и тогда вместе с лепидодендронами, сигилляриями, папоротниками и кордаитами образовывали леса, которые дали залежи каменного угля.
По внешнему виду и строению каламиты напоминали современные хвощи, однако отличались от них - были деревьями, достигали в высоту 8-10 м и даже до 20 м. Среди них были как равноспоровые, так и разноспоровые виды.
Семейство хвощовых включает один род хвощ (Equisetum) и 25 видов. В Республике Беларусь растет 8 видов хвоща. Они встречаются на болотах (E. palustre, E. fluviatile), в лесах (E. sylvaticum), в кустарниках (E. hyemale), на лугах, на полях (E. pratense, E. arvense) и др.
Современные хвощи - небольшие травянистые растения 80-100 см высотой, 2-5 мм толщиной. Тропический южноамериканский E. giganteum достигает 10-12 м длины и является лианой.
Хвощ состоит из горизонтально расположенного в почве корневища, от узлов которого отходят тонкие корни и вверх поднимаются надземные побеги.
Стебель хвоща членистый, ребристый, состоит из узлов и междоузлий. Междоузлия полые в середине, узлы заполнены паренхиматической тканью.
Листья хвощей чешуевидные, бурые, коричневые, лишены хлорофилла, срослись в нижней части в трубчатое влагалище, прикрепленное к узлу. Вследствие редукции листьев функцию ассимиляции выполняют зеленые побеги и стебель. Ветви расположены мутовками, пробивая влагалище сросшихся листьев.
На поперечном разрезе стебель имеет следующее строение. Снаружи стебель неровный, имеются возвышенные участки (ребрышки), чередующиеся с ложбинками. Снаружи стебель покрыт однослойным эпидермисом, пропитанным кремнеземом, что придает ему прочность. Внутрь от эпидермиса расположена кора и кольцо мелких, изолированных проводящих пучков коллатерального типа с каринальными (от лат. сarina - киль, гребень) каналами. В центре стебля находится полость на месте разрушений сердцевины. Под ребрышками расположены участки механической ткани, а под ложбинками - а с с и м и л я ц и о н н а я т к а н ь и валекулярные (от лат. vallis - долина, ложбинка) полости. Под механической тканью (под ребрышками) находятся проводящие пучки коллатерального типа, закрытые, без камбия. В эпидермисе, над ассимиляционной тканью имеются устьица.
Спороносные колоски у хвощей возникают по одному на верхушке главного побега, иногда и боковых ветвей. У большинства видов спороносный побег зеленый. Надземные побеги у одних видов могут совмещать две функции - спороносную и вегетативную. Так, у хвоща болотного (E. palustre) и приречного, или топеного (E. fluviatila), вегетативные и спороносные побеги возникают одновременно и не отличаются морфологически друг от друга. Только в середине лета на некоторых зеленых побегах формируются стробилы. У других видов наблюдается разделение функций побегов. Так, у хвоща лесного (E. silvaticum) и хвоща лугового (E. pratense) весной одновременно с вегетативными побегами развиваются неветвящиеся бесцветные или розоватые спороносные побеги. Но после спороношения они зеленеют, ветвятся и не отличаются от вегетативных побегов. У некоторых видов диморфия побегов проявляется очень четко.
У хвоща полевого побеги двух типов. Весной от корневища вырастают бурые, спороносные побеги, несущие один колосок. Колосок хвоща состоит из многочисленных спорангиофоров, собранных мутовками на его оси. Спорангиофоры состоят из ножки и щитковидного шестигранного диска. На нижней стороне диска, вокруг ножки, располагается 5-13 мешковидных спорангиев. В спорангиях образуется большое количество одинаковых спор (равноспоровость). Спора имеет три оболочки: эндоспорий, экзоспорий и внешний слой оболочки, который при созревании растрескивается с образованием вокруг споры двух гигроскопичных лент, получивших название г а п т е р, которые прикрепляются в центре к споре. В сухую погоду они, как пружинки, раскручиваются и способствуют разрыхлению спор. При этом гаптеры соседних спор цепляются друг за друга. В результате из спорангиев высыпаются рыхлые комочки спор, легко разносимые ветром. Заросток хвоща имеет вид зеленой пластинки, а в загущенных посевах или в воде - зеленую нить. Однослойная пластинка, разрастаясь, превращается в многослойную распростертую подушку с ризоидами на нижней стороне. На верхней стороне подушка разхвиваются вертикальные пластинчатые лопасти, на которых формируются половые органы. Размеры гаметофитов у разных видов варьируют от 1 мм до 2-3 см. В пределах вида мужские гаметофиты мельче женских.
Некоторые виды хвоща имеют физиологическую разноспоровость.
В лучших условиях увлажнения, освещения из спор развиваются более крупные заростки (женские), в худших - мелкие заростки (мужские).
Антеридии хвощей погружены в ткань заростка. В них развивается до 100 многожгутиковых сперматозоидов. Архегонии шейкой возвышаются над заростком. Оплодотворение происходит в сырую погоду. Зародыш подвеска не образует и состоит из стебелька, 2-3 листочков и корешка.
После выпадения спор из колосовидного стробика хвоща полевого спороносный побег отмирает. Из корневища вырастают новые - зеленые, сильно ветвистые летние побеги.
П р а к т и ч е с к о е з н а ч е н и е хвощей невелико. Стебли, содержат кремнезем и потому употребляются для чистки металлической посуды и полировки дерева. Клубеньки на корневище хвоща полевого иногда идут в пищу (содержит крахмал). Некоторые из хвощей (хвощ полевой, хвощ луговой) - сорняки. Некоторые ядовиты (хвощ болотный).
В Красную книгу Республики Беларусь внесены редкие реликтовые виды - хвощ большой (Equisetum telmateia) и хвощ пестрый (E. variegatum) .
Литература:
1. Еленевский А.Г., Соловьева М.П., Тихомиров В.Н. Ботаника высших, или наземных растений: Учеб. для студ. высш. пед. учеб. заведений.-М.: Издательский центр "Академия", 2000.-432 с.
2. Комарницкий Н.А., Кудряшов Л.В., Уранов А.А. Ботаника: Систематика растений.- 7-е изд., перераб. - М.: Просвещение, 1975. - 608 с.
3. Чырвоная кніга Рэспублікі Беларусь: Рэдкія і тыя, што знаходзяцца пад пагрозай знікнення віды жывелін і раслін / Гал. рэдкал.: А.М. Дарафееў (старш.) і інш. - Мн.: БелЭн., 1993. - 560 с.
Представьте себе такую картину. Вышли вы вечером на крыльцо дома, подняли голову вверх и заметили в ночном небе небольшую светящуюся точку. Эта точка по мере приближения к поверхности Земли всё увеличивалась и увеличивалась, пока вы не осознали, что размеры этой точки не меньше города Москвы. Дальше оглушительный гул, взрыв, землетрясения и пыль, которая закроет тёмным покрывалом Землю от солнечных лучей на долгие годы. Такие катаклизмы в истории Земли происходили не раз, именно с ними учёные связывают гибель динозавров и других организмов нашей планеты. Environmentalgraffiti.com, в дополнение к рейтингам , и , опубликовала рейтинг самых больших «шрамов Земли», вызванных падением астероидов.
10. Кратер Barringer в Аризоне, США
Приблизительно 49 000 лет назад железно-никелевый метеорит диаметром около 46 метров и массой около 300 000 тонн летевший со скоростью около 18 километров в секунду «приземлился» в Аризоне. Сила взрыва была эквивалентна силе взрыва 20 миллионов тонн тротила, от такого чудовищного взрыва образовался кратер диаметром 1,2 километра (в 26 раз больше диаметра самого метеорита), глубиной 75 метров и валом, опоясывающим воронку, высотой 45 метров. Кратер носит имя горного инженера Daniel Barringer, который его впервые открыл. Этот кратер до сих пор является собственностью его семьи. Этот шрам на лице нашей планеты ещё известен под названиями «Кратер Метеора», «Енот Бьютт» и «Каньон Дьявола».
9. Bosumtwi, Гана
Источник: .
В 30 километрах на юго-восток от Кумаси на идеально-ровном Южно-африканском щите находится единственное озеро страны Bosumtwi. Это озеро было образовано падением метеорита 1,3 миллиона лет назад, который оставил после себя кратер диаметром 10,5 километров. Кратер постепенно заполнился водой и превратился в озеро, окружённое пышной тропической растительностью. Для проживающего здесь африканского племени Ашанти это озеро является священным. Согласно их поверьям именно здесь души умерших встречаются с богом Туи.
8. Глубокий Залив, Канада
Источник: www.ersi.ca
Этот 13 километровый кратер, тоже заполнен водой, расположен недалеко от Оленьего озера в Канаде. Этот метеорит упал на Землю приблизительно 100 – 140 миллионов лет назад.
7. Aorounga, кратер в Чад
Метеорит, вызвавший появление кратера Aorounga, «совершил посадку» в пустыне Сахара северного Чада 2-300 миллионов лет назад. Такие метеориты падают на нашу планету с периодичностью раз в миллион лет. Диаметр метеорита составлял приблизительно 1,6 километров. Его падение вызвало появление на теле нашей планеты кратера диаметром 17 километров. Самое удивительно то, что кратер окружают кольцеобразные образования. Учёные предполагают, что они образованы обломками метеорита, образовавшимися при прохождении астероида сквозь плотные слои атмосферы.
6. Gosses Bluff, Австралия
Источник: , ,
Приблизительно 142 миллиона лет назад астероид или комета диаметром 22 километра на скорости 40 километров в секунду «поцеловал» нашу планету, практически в центр материка Австралия. Взрыв был эквивалентен взрыву 22 000 мегатонн тротила. От взрыва чудовищной силы образовалась воронка диаметром 24 километра и глубиной 5 километров.
5. Озеро Мистастин, Канада
Источник:
Озеро Мистастин на полуострове Лабрадор в Канаде не что иное как след от падения метеорита 38 миллионов лет назад. Падение метеорита вызвало образование кратера диаметром 28 километров, которое впоследствии заполнилось водой. Посредине озера, образованного падением метеорита, есть остров, который,по-видимому, образовался вследствие неоднородной структуры упавшего метеорита.
4. Озёра Чистая вода, Канада
Два круглых кратера на Канадском щите, сейчас тоже заполненных водой, образовались при столкновении метеорита с Землёй около 290 миллионов лет назад. Кратеры располагаются в Квебеке на восточном берегу Гудзонова залива. Диаметр западного кратера составляет 32 километра, восточного – 22 километра. Эти кратеры благодаря своим «рваным» краям, образующим большое количество островов, пользуются большой популярностью у туристов.
3. Каракуль, Таджикистан, СНГ
Всесильный Космос не обделил своим вниманием и СНГ. На высоте 3 900 метров над уровнем моря, в горах Памира Таджикистана, недалеко от границы с Китаем располагается озеро. Это озеро образовалось в астероидном кратере диаметром 45 километров. Падение произошло приблизительно 5 миллионов лет назад.
2. Manicouagan, Канада







