ЯДЕРНАЯ СТРУКТУРА АТОМА
Альфа-частицы . В 1896 году французским физиком Беккерелем было открыто явление радиоактивности. После этого начался быстрый прогресс в изучении строения атома. Этому, прежде всего способствовало то, что в руках физиков оказался очень эффективный инструмент исследования атомной структуры – α -частица. С помощью α -частиц, испускаемых естественными радиоактивными веществами, были сделаны важнейшие открытия: установлена ядерная структура атома, получены первые ядерные реакции, обнаружено явление искусственной радиоактивности и, наконец, найден нейтрон, сыгравший важную роль как при объяснении строения атомного ядра, так и при открытии процесса деления тяжелых ядер.
Альфа-частицы – это движущиеся с большой скоростью ядра гелия. Измерения скоростей α- частиц естественных излучателей по отклонению в электрическом и магнитном полях дали величину скорости (1,5-2).10 7 м/с, что соответствует кинетической энергии 4,5-8 МэВ (1 МэВ=1,6.10 -13 Дж). Такие частицы движутся в веществе прямолинейно, быстро теряют энергию на ионизацию атомов и после остановки превращаются в нейтральные атомы гелия.
Рассеяние альфа-частиц. Опыты Резерфорда. Изучая прохождение коллимированного пучка альфа-частиц через тонкую металлическую фольгу, английский физик Резерфорд обратил внимание на размытие изображения пучка частиц на регистраторе – фотопластинке. Резерфорд объяснил это размытие рассеянием альфа-частиц. Детальное изучение рассеяния альфа-частиц показало, что в редких случаях они рассеиваются на большие углы иногда превышающие 90 0 , что соответствует отбрасыванию быстродвижущихся частиц в обратном направлении. Такие случаи рассеяния в рамках модели Томпсона объяснить нельзя.
Тяжелая альфа-частица в одном акте столкновения может быть отброшена назад только при взаимодействии с частицей большей массы, превосходящей массу альфа-частицы. Электроны не могут быть такими частицами. Кроме того, рассеяние в обратном направлении предполагает сильное торможение альфа-частицы, т.е. энергия взаимодействия должна быть порядка кинетической энергии альфа-частицы. Энергия же электростатического взаимодействия альфа-частицы с атомом Томпсона, имеющим положительный заряд, распределенный в объеме или на поверхности атома радиусом 10 -8 см и равный в единицах элементарного заряда примерно половине атомной массы, много меньше этой величины. Результаты опыта можно объяснить, если расстояние от альфа частицы до центра положительного электрического заряда порядка 10 -12 см. Такое расстояние в 10000 раз меньше радиуса атома, а радиус положительного заряда должен быть еще меньше. Предположение о малом объеме рассеивающего центра согласуется с очень малым количеством случаев рассеяния на большие углы.
Для объяснения результатов своих наблюдений по рассеянию альфа-частиц Резерфорд предложил ядерную модель атома . Согласно этой модели в центре атома находится ядро, занимающее очень малый объем, содержащее почти всю массу атома и несущее положительный электрический заряд. Основной объем атома занят движущимися электронами, число которых равно числу элементарных положительных зарядов ядра, т.к. атом в целом нейтрален.
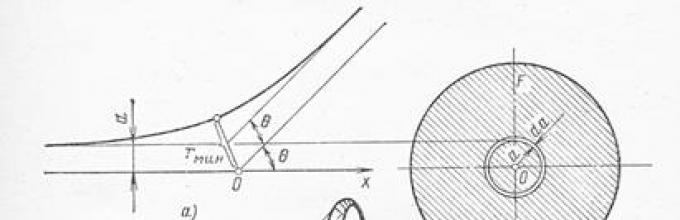
Теория рассеяния альфа-частиц. Чтобы обосновать предположение о ядерной структуре атома и доказать, что рассеяние альфа-частиц происходит в результате кулоновского взаимодействия с ядром, Резерфорд развил теорию рассеяния альфа-частиц точечными электрическими зарядами с большой массой и получил зависимость между углом рассеяния θ и числом частиц, рассеянных на угол θ . Если альфа-частица движется в направлении точечного заряда Ze , где Z – число элементарных зарядов, и при этом ее начальная траектория отстоит от оси, проходящей через рассеивающий центр, на расстоянии а (рис.1.1), то на основании закона Кулона методами классической механики можно вычислить угол θ , на который отклонится альфа-частица вследствие электростатического отталкивания одноименных электрических зарядов:
где M и v – масса и скорость альфа-частицы; 2e – ее заряд; ε 0 – электрическая постоянная, равная 8,85.10 -12 Ф/м.

Рис.1.1. Рассеяние альфа-частицы электрическим полем атомного ядра:
а) – схема рассеяния в плоскости траектории частицы; б) – кольцо, из которого происходит рассеяние под углом θ ; в) – схема рассеяния в конический телесный угол под угломθ к оси.
Доля частиц dn/n 0 , имеющих параметр удара а , от полного числа n 0 , падающих на мишень, равна доле элементарной площадки 2πada на полной площади F поперечного сечения пучка альфа-частиц (рис. 1.1, б). Если на площади F имеется не один, а N F рассеивающих центров, то соответствующая доля возрастет в N F раз и отнесенная к единице а, составит:
![]() , (1.2)
, (1.2)
где N 1 – число рассеивающих центров на единице площади мишени.
Учитывая, что dΩ=2π sinθ dθ, можно получить долю частиц, рассеянных в единицу конического телесного угла под углом θ к оси, как :
 (1.3)
(1.3)
Экспериментальная проверка полностью подтвердила последнюю зависимость при рассеянии веществом альфа-частиц. Строгое выполнение закона 1/sin 4
свидетельствует о том, что за рассеяние ответственны только электрические силы и что геометрические размеры электрических зарядов обоих тел по крайней мере меньше наименьшего расстояния в акте рассеяния r мин
. Расстояние r мин
тем меньше, чем больше угол рассеяния θ
. При θ
=π () оно самое малое и определяется условием ![]() , которое соответствует случаю обращения всей кинетической энергии альфа-частицы в потенциальную энергию отталкивания одноименных зарядов.
, которое соответствует случаю обращения всей кинетической энергии альфа-частицы в потенциальную энергию отталкивания одноименных зарядов.
По результатам обработки результатов экспериментов, исходя из различных в то время оценок величины заряда ядер Z , Резерфорд оценил радиус ядра величиной порядка 10 -12 см.
Атом Резерфорда-Бора. С открытием атомного ядра возникла необходимость объяснения устойчивости атома. С точки зрения классической электродинамики атом Резерфорда не может существовать длительно. Поскольку разноименные заряды притягиваются, электроны могут находится на определенном расстоянии от ядра только при условии движения вокруг ядра. Однако движение по замкнутой траектории является движением с ускорением, а движущийся с ускорением электрический заряд излучает энергию в окружающее пространство. За ничтожно малое время любой атом должен излучить энергию движения электронов и уменьшиться до размера ядра.
Первую стационарную модель атома предложил датский физик Нильс Бор в 1913 году. Бор связал устойчивость атомов с квантовой природой излучения. Гипотеза квантов энергии, выдвинутая немецким физиком Планком в 1900 году для объяснения спектра излучения абсолютно черного тела, утверждала, что микроскопические системы способны излучать энергию лишь определенными порциями – квантами с частотой v , пропорциональной энергии кванта E:
где h – универсальная постоянная Планка, равная 6,62.10 -24 Дж.с.
Бор предположил, что энергия атомного электрона в кулоновском поле ядра не меняется непрерывно, а принимает ряд устойчивых дискретных значений, которым соответствуют стационарные орбиты электронов. При движении по таким орбитам электрон не излучает энергию. Излучение атома происходит только при переходе электрона с орбиты с более высоким значением энергии на другую стационарную орбиту. Это излучение характеризуется единственным значением частоты, пропорциональной разности энергий орбит:
hv=Е нач - Е кон
Условие стационарности орбиты – это равенство механического момента количества движения электрона целому кратному числу h/2π :
mvr n = n ,
где mv – модуль импульса электрона;
r n – радиус n -ой стационарной орбиты;
n – любое целое число.
Введенное Бором условие квантования круговых орбит позволило рассчитать спектр атома водорода и вычислить спектроскопическую константу Ридберга для атома водорода. Система уровней одноэлектронного атома и радиусы стационарных орбит можно определить из последнего соотношения и закона Кулона:
![]() ; (1.4)
; (1.4)
Расчет по этим формулам при n=1 и Z=1 дает радиус наименьшей стационарной орбиты электрона в атоме водорода или первый боровский радиус:
![]() . (1.6)
. (1.6)
Движение электрона по орбите можно представить как замкнутый электрический ток и вычислить создаваемый им магнитный момент. Для первой орбиты водорода он называется магнетоном Бора и равен:
![]() (1.7)
(1.7)
Магнитный момент обратно пропорционален массе частицы, но для частиц данного сорта, например электронов, имеет смысл единицы. Характерно, что как раз этой единице равен собственный момент электрона, связанный с его спином.
Ядерная модель атома с электронами на устойчивых орбитах называется планетарной моделью Резерфорда-Бора. Она не приводит к верным количественным результатам в приложении к атомам с более, чем одним электроном, но зато очень удобна при качественной интерпретации атомных явлений. Точную теорию атома дает квантовая механика.
Дискретная природа микромира. Открытие атомного строения вещества оказалось первым шагом на пути открытий дискретной природы микромира. Не только массы и электрические заряды микротел дискретны, но и динамические величины, описывающие состояния микросистем, такие, как энергия, момент количества движения, также дискретны и характеризуются скачкообразным изменением своих численных значений.
В процессе эволюции претерпевали ряд изменений. Появлению новых органелл предшествовали преобразования в атмосфере и литосфере молодой планеты. Одним из значительных приобретений стало клеточное ядро. Эукариотические организмы получили, благодаря наличию обособленных органелл, существенные преимущества перед прокариотами и быстро стали доминировать.
Клеточное ядро, строение и функции которого несколько отличаются в разных тканях и органах, позволило повысить качество биосинтеза РНК и передачу наследственной информации.
Происхождение
На сегодняшний день есть две основные гипотезы об образовании эукариотической клетки. Согласно симбиотической теории органеллы (например, жгутики или митохондрии) когда-то были отдельными прокариотическими организмами. Предки современных эукариот поглотили их. В результате образовался симбиотический организм.
Ядро при этом сформировалось в результате выпячивания внутрь участка цитоплазматической было необходимым приобретением на пути освоения клеткой нового способа питания, фагоцитоза. Захват пищи сопровождался повышением степени подвижности цитоплазмы. Генофоры, представлявшие собой генетический материал прокариотической клетки и прикреплявшиеся к стенкам, попадали в зону сильного «течения» и нуждались в защите. В результате и образовалось глубокое впячивание участка мембраны, содержавшего прикрепленные генофоры. В пользу этой гипотезы свидетельствует тот факт, что оболочка ядра неразрывно связана с цитоплазматической мембраной клетки.
Существует и другая версия развития событий. Согласно вирусной гипотезе происхождения ядра, оно сформировалось в результате заражения клетки древней археи. В нее внедрился ДНК-вирус и постепенно получил полный контроль над жизненными процессами. Ученые, считающие эту теорию более правильной, приводят массу доводов в ее пользу. Однако на сегодняшний день нет исчерпывающего доказательства ни для одной из существующих гипотез.
Одно или несколько
Большая часть клеток современных эукариот имеет ядро. Подавляющее их число содержит только одну подобную органеллу. Существуют, однако, и клетки, которые утратили ядро по причине некоторых функциональных особенностей. К ним относятся, например, эритроциты. Встречаются и клетки с двумя (инфузории) и даже несколькими ядрами.
Структура клеточного ядра

Вне зависимости от особенностей организма, строение ядра характеризуется набором типичных органелл. От внутреннего пространства клетки оно отгорожено двойной мембраной. Внутренние и внешние ее прослойки в некоторых местах сливаются, образуя поры. Их функция заключается в обмене веществ между цитоплазмой и ядром.
Пространство органеллы заполнено кариоплазмой, также называемой ядерным соком или нуклеоплазмой. В ней размещается хроматин и ядрышко. Иногда последний из названных органоид клеточного ядра присутствует не в единственном экземпляре. У некоторых же организмов ядрышки, наоборот, отсутствуют.
Мембрана
Ядерная оболочка образована липидами и состоит из двух слоев: наружного и внутреннего. По сути, это та же клеточная мембрана. Ядро сообщается с каналами эндоплазматической сети через перинуклеарное пространство, полость, образованную двумя слоями оболочки.
Наружная и внутренняя мембрана имеют свои особенности в строении, однако в целом довольно похожи.
Ближайший к цитоплазме
Наружный слой переходит в мембрану эндоплазматической сети. Ее основное отличие от последней — значительно более высокая концентрация белков в структуре. Мембрана, непосредственно контактирующая с цитоплазмой клетки, покрыта слоем рибосом с наружной стороны. С внутренней мембраной она соединяется многочисленными порами, представляющими собой довольно крупные белковые комплексы.
Внутренний слой
Обращенная в клеточное ядро мембрана, в отличие от наружной, гладкая, не покрытая рибосомами. Она ограничивает кариоплазму. Характерная особенность внутренней мембраны — слой ядерной ламины, выстилающий ее со стороны, соприкасающейся с нуклеоплазмой. Эта специфическая белковая структура поддерживает форму оболочки, участвует в регуляции экспрессии генов, а также способствует прикреплению хроматина к мембране ядра.
Обмен веществ
Взаимодействие ядра и цитоплазмы осуществляется через Они представляют собой довольно сложные структуры, образованные 30 белками. Количество пор на одном ядре может быть разным. Он зависит от типа клетки, органа и организма. Так, у человека клеточное ядро может иметь от 3 до 5 тысяч пор, у некоторых лягушек оно доходит до 50 000.

Главная функция пор — обмен веществ между ядром и остальным пространством клетки. Некоторые молекулы проникают сквозь поры пассивно, без дополнительных затрат энергии. Они обладают небольшими размерами. Транспортировка крупных молекул и надмолекулярных комплексов требует расхода определенного количества энергии.
Из кариоплазмы в клетку попадают синтезируемые в ядре молекулы РНК. В обратном направлении транспортируются белки, необходимые для внутриядерных процессов.
Нуклеоплазма
Строение ядерного сока меняется в зависимости от состояния клетки. Их два — стационарное и возникающее в период деления. Первое характерно для интерфазы (время между делениями). При этом ядерный сок отличается равномерным распределением нуклеиновых кислот и неструктурированными молекулами ДНК. В этот период наследственный материал существует в виде хроматина. Деление клеточного ядра сопровождается преобразованием хроматина в хромосомы. В это время изменяется строение кариоплазмы: генетический материал приобретает определенную структуру, ядерная оболочка разрушается, и кариоплазма смешивается с цитоплазмой.
Хромосомы

Основные функции нуклеопротеидных структур преобразованного на время деления хроматина — хранение, реализация и передача наследственной информации, которую содержит клеточное ядро. Хромосомы характеризуются определенной формой: делятся на части или плечи первичной перетяжкой, также называемой целомерой. По ее расположению выделяют три типа хромосом:
- палочкообразные или акроцентрические: для них характерно размещение целомеры практически на конце, одно плечо получается очень маленьким;
- разноплечие или субметацентрические обладают плечами неравной длины;
- равноплечие или метацентрические.
Набор хромосом в клетке называется кариотипом. У каждого вида он фиксирован. При этом разные клетки одного организма могут содержать диплоидный (двойной) или гаплоидный (одинарный) набор. Первый вариант характерен для соматических клеток, в основном составляющих тело. Гаплоидный набор — привилегия половых клеток. Соматические клетки человека содержат 46 хромосом, половые — 23.
Хромосомы диплоидного набора составляют пары. Одинаковые нуклеопротеидные структуры, входящие в пару, называются аллельными. Они имеют одинаковое строение и выполняют одни и те же функции.
Структурной единицей хромосом является ген. Он представляет собой участок молекулы ДНК, кодирующий определенный белок.
Ядрышко

Клеточное ядро обладает еще одним органоидом — это ядрышко. Оно не отделяется от кариоплазмы мембраной, но при этом его легко заметить во время изучения клетки с помощью микроскопа. Некоторые ядра могут иметь несколько ядрышек. Существуют и такие, в которых подобные органоиды отсутствуют совсем.
По форме ядрышко напоминает сферу, имеет достаточно небольшие размеры. В его состав входят различные белки. Основная функция ядрышка — синтез рибосомных РНК и самих рибосом. Они необходимы для создания полипептидных цепей. Ядрышки образуются вокруг специальных участков генома. Они получили название ядрышковых организаторов. Здесь содержатся гены рибосомной РНК. Ядрышко, кроме прочего, является местом с наибольшей концентрацией белка в клетке. Часть белков необходима для выполнения функций органоида.
В составе ядрышка выделяют два компонента: гранулярный и фибриллярный. Первый представляет собой созревающие субъединицы рибосом. В фибриллярном центре осуществляется Гранулярный компонент окружает фибриллярный, расположенный в центре ядрышка.
Клеточное ядро и его функции
Роль, которую играет ядро, неразрывно связана с его строением. Внутренние структуры органоида совместно реализуют важнейшие процессы в клетке. Здесь размещается генетическая информация, которая определяет строение и функции клетки. Ядро отвечает за хранение и передачу наследственной информации, осуществляющееся во время митоза и мейоза. В первом случае дочерняя клетка получает идентичный материнскому набор генов. В результате мейоза образуются половые клетки с гаплоидным набором хромосом.

Другая не менее важная функция ядра — регуляция внутриклеточных процессов. Она осуществляется в результате контроля синтеза белков, отвечающих за строение и функционирование клеточных элементов.
Влияние на белковый синтез имеет еще одно выражение. Ядро, контролируя процессы внутри клетки, объединяет все ее органоиды в единую систему с отлаженным механизмом работы. Сбои в нем приводят, как правило, к гибели клетки.
Наконец, ядро является местом синтеза субъединиц рибосом, которые отвечают за образование все того же белка из аминокислот. Рибосомы незаменимы в процессе транскрипции.

Представляет собой более совершенную структуру, чем прокариотическая. Появление органоидов с собственной мембраной позволило повысить эффективность внутриклеточных процессов. Формирование ядра, окруженного двойной липидной оболочкой, играло в этой эволюции очень важную роль. Защита наследственной информации мембраной позволила освоить древним одноклеточным организмам новые способы жизнедеятельности. Среди них был фагоцитоз, который по одной из версий привел к появлению симбиотического организма, позже ставшего прародителем современной эукариотической клетки со всеми характерными для нее органоидами. Клеточное ядро, строение и функции некоторых новых структур позволили задействовать кислород в метаболизме. Следствием этого стало кардинальное изменение в биосфере Земли, была заложена основа для формирования и развития многоклеточных организмов. Сегодня эукариотические организмы, к которым относится и человек, доминируют на планете, и ничто не предвещает изменений в этом плане.
Введение
Глава 1. Структура и химия клеточного ядра. Открытие ядра. Роберт Броун
1.1. Интерфазное ядро
1.2. Работы флемминга
1.3. Ядрышки
1.4. Ядерная мембрана
1.5. Кариоплазма
1.6. Хроматин
Глава 2. Клеточное ядро - центр управления жизнедеятельностью клетки
2.1. Ядро - незаменимый компонент клетки
2.2. Функциональная структура ядра
2.3. Роль ядерных структур в жизнедеятельности клетки
2.4. Ведущее значение днк
Список литературы
ВВЕДЕНИЕ
Клеточное ядро - центр управления жизнедеятельностью клетки. Из общей схемы белкового синтеза можно видеть, что начальным пунктом, с которого начинается поток информации для биосинтеза белков в клетке, является ДНК. Следовательно, именно ДНК содержит ту первичную запись информации, которая должна сохраняться и воспроизводиться от клетки к клетке, из поколения в поколение. Кратко касаясь вопроса о месте хранения генетической информации, т. е. о локализации ДНК в клетке, можно сказать следующее. Уже давно известно, что, в отличие от всех прочих компонентов синтезирующего белок аппарата, универсально распределенных по всем частям живой клетки, ДНК имеет особую, весьма ограниченную локализацию: местом ее нахождения в клетках высших (эукариотических) организмов является клеточное ядро.
У низших (прокариотических) организмов, не имеющих оформленного клеточного ядра, - бактерий и сине-зеленых водорослей, - ДНК также отделена от остальной части протоплазмы одним или несколькими компактными нуклеоидными образованиями. В полном соответствии с этим ядро эукариотов или нуклеоид прокариотов издавна рассматриваются как вместилище генов, как уникальный клеточный органоид, контролирующий реализацию наследственных признаков организмов и их передачу в поколениях. Генетические данные о «единоначалии» ядра в клетке всегда непосредственно объединялись с биохимическими данными об уникальной локализации ДНК в ядре.
1. СТРУКТУРА И ХИМИЯ КЛЕТОЧНОГО ЯДРА. ОТКРЫТИЕ ЯДРА. РОБЕРТ БРОУН
Термин «ядро» впервые был применен Броуном в 1833 г. для обозначения шаровидных постоянных структур в клетках растений. В 1831-1833 гг., шотландский путешественник и физик (открывший «броуновское движение») Роберт Броун (1773-1858) обнаружил ядро в растительных клетках. Он дал ему название «Nucleus», или «Areola». Первый термин стал общепринятым и сохранился по настоящее время, второй же распространения не получил и забыт. Весьма важно, что Броун настаивал на постоянном наличии ядра во всех живых клетках.
Роль и значение клеточного ядра не были в то время известны. Полагали, что оно представляет собой «конденсированную в комочек слизь, а возможно, и запасное питательное вещество». Позднее такую же структуру описали во всех клетках высших организмов. Говоря о клеточном ядре, мы имеем в виду собственно ядра эукариотических клеток. Их ядра построены сложным образом и довольно резко отличаются от «ядерных» образований, нуклеоидов прокариотических организмов. У последних в состав нуклеоидов (ядроподобных структур) входит одиночная, кольцевая молекула ДНК, практически лишенная белков. Иногда такую молекулу ДНК бактериальных клеток называют бактериальной хромосомой, или генофором (носителем генов).
Бактериальная хромосома не отделена мембранами от основной цитоплазмы, однако собрана в компактную, ядерную зону, нуклеоид, который можно видеть в световом микроскопе после специальных окрасок или же в электронном микроскопе. Анализируя структуру и химию клеточного ядра, мы будем опираться на данные, касающиеся ядер эукариотических клеток, постоянно сравнивая их с ядрами прокариотов. Клеточное ядро, обычно одно на клетку (есть примеры многоядерных клеток), состоит из ядерной оболочки, отделяющей его от цитоплазмы, хроматина, ядрышка и кариоплазмы или ядерного сока. Эти четыре основных компонента встречаются практически во всех неделящихся клетках эукариотических одно- или многоклеточных организмов.
1.2. РАБОТЫ ФЛЕММИНГА
До некоторых пор роль ядра в клеточном делении оставалась неопределенной. Это, вероятно, было связано с трудностью наблюдения за ним. В живой клетке ядро, как правило, можно видеть только при значительном увеличении обычного светового микроскопа. Ядро, находящееся в процессе деления, наблюдать еще труднее. Анилиновые красители окрашивают ядро, цитоплазму и клеточную оболочку по-разному и, следовательно, облегчают узнавание этих структур.
Анилиновые красители синтезируются искусственно, и методика их получения не была известна до середины XIX в. Естественные красители, которые биологи использовали раньше, не всегда окрашивали ядра достаточно хорошо, чтобы их можно было отличить от остальных частей клетки. И вновь дальнейший прогресс зависел от развития подходящих для проведения исследований методов. В то время не было недостатка в хороших микроскопах, но не было известно, как обрабатывать клетки, чтобы увидеть как можно больше клеточных структур. Следует отметить, что никто не знал, будут ли анилиновые красители для этой цели лучше, чем естественные.
Когда в 1860-х гг. химики получили анилиновые красители, кто-то просто наугад попытался использовать их для окрашивания тонких срезов растительных и животных тканей. В 1879 г. немецкий биолог Вальтер Флемминг использовал различные анилиновые красители и ахроматические линзы. Обработав клетки красителями и изучая их под микроскопом с ахроматическими линзами, он проследил за поведением ядра в процессе клеточного деления. В его книге «Клеточное вещество, ядро и клеточное деление» описаны результаты наблюдений над клеточным делением, причем описания очень близки к современным.
Поскольку хромосомы похожи на нити, Флемминг решил назвать этот процесс митозом (греческое слово, что в переводе значит «нить»). Строго говоря, митоз относится только к процессу ядерного удвоения. Образования клеточной пластинки в растительных клетках и клеточной бороздки в животных клетках являются делениями цитоплазмы.
Было бы неправильным считать, что Флемминг - единственный первооткрыватель явления митоза. Понимание всей последовательности процесса митоза зависело от многих ученых, работавших над этой проблемой все предыдущие годы. Одна из основных трудностей исследования событий, происходящих в клетке, состояла в том, что клетки погибали в процессе окрашивания. Это означает, что клетка изучается только после того, как жизнедеятельность в ней прекращена. По этой «остановленной в движении» картине Флемминг и другие исследователи воссоздали то, что происходит в живых клетках. Это примерно то же, что воссоздать работу фабрики по серии моментальных снимков, взятых в различные интервалы времени. По существу, это и было сделано Флеммингом. Другие ученые, основываясь на работе Флеминга, в конце концов выявили связь хромосом с наследственностью и эволюцией.
Именно так развивается наука: успех зависит не от случайных открытий ученых-«гигантов», а от кропотливой работы большого отряда ученых. В световом, а также в фазово-контрастном микроскопах ядро обычно представляется оптически гомогенным: видны лишь оболочка и одно или несколько ядрышек внутри. Иногда обнаруживаются также гранулы и небольшие глыбки. Реже в неделящихся живых клетках удается наблюдать хромосомы. Тонкая хроматиновая сеть отчетливо выявляется лишь после фиксации и окрашивания клетки основными красителями.
Исследования ядра на фиксированных и окрашенных препаратах показали, что его микроскопическое изображение почти не зависит от метода изготовления препаратов. Лучше всего тонкая структура ядра сохраняется при фиксации четырехокисью осмия. Другие общепринятые фиксаторы позволяют различать на препарате ядерную оболочку, ядрышко, хроматиновые структуры в виде глыбок и нитей и неокрашенную массу между ними - нуклеоплазму.
Хроматиновые структуры расположены в более жидкой ахроматической среде, они могут быть плотными или рыхлыми, пузыревидными. У некоторых объектов хроматин после фиксации не образует явно выраженной ядерной сети, а концентрируется в ядре в виде крупных глыбок, названных хромоцентрами, или прохромосомами. В ядрах подобного типа весь хроматин сосредоточен в хромоцентрах.
1.3. ЯДРЫШКИ
Согласно электронно-микроскопическим исследованиям, ядрышки лишены какой-либо мембраны. Вещество их в основном состоит из субмикроскопических нитей и нуклеоплазмы. Ядрышки можно наблюдать, применяя специальные методы окрашивания, а также в ядрах некоторых живых клеток при использовании фазово-контрастного микроскопа или темнопольного конденсора.
На электронных микрофотографиях в ядрышках нередко видны две зоны: центральная - гомогенная и периферическая - построенная из гранулированных нитей. Эти гранулы напоминают рибосомы, но отличаются от них меньшей плотностью и величиной. Ядрышки богаты белками (80-85 %) и РНК (около 15 %) и служат активными центрами синтеза рибосомальной РНК. В соответствии с этим главной составной частью ядрышка является ядрышковая ДНК, которая принадлежит организатору ядрышек одной из хромосом.
содержание РНК заметно колеблется, в зависимости от интенсивности обмена веществ в ядре и цитоплазме. Ядрышки не присутствуют в ядре постоянно: они возникают в средней телофазе митоза и исчезают в конце профазы. Полагают, что по мере затухания синтеза РНК в средней профазе происходят разрыхление ядрышка и выход в цитоплазму образовавшихся в нуклеоплазме субчастиц рибосом. При исчезновении ядрышка во время митоза его белки, ДНК и РНК, становятся основой матрикса хромосом, а в дальнейшем из материала старого ядрышка формируется новое.
Установлена связь ядрышек с хромосомами, имеющими спутников, поэтому число ядрышек соответствует числу спутничных хромосом. Нуклеолонемы сохраняются на протяжении всего цикла клеточного деления и в телофазе переходят от хромосом к новому ядрышку.
1.4. ЯДЕРНАЯ МЕМБРАНА
Неделящееся клеточное ядро заключено в плотную и упругую оболочку, которая растворяется и вновь восстанавливается в процессе деления клетки. Это образование отчетливо видно лишь на некоторых объектах, например у гигантских ядер слизевых клеток алоэ толщина мембраны достигает 1 мкм. В световом микроскопе структуру ядерной оболочки удается наблюдать лишь у плазмолизированных клеток, фиксированных и окрашенных.
Детальное изучение ядерной мембраны стало возможным с появлением электронной микроскопии. Исследования показали, что наличие ядерной оболочки характерно для всех эукариотических клеток. Она состоит из двух элементарных мембран толщиной 6-8 нм каждая - внешней и внутренней, между которыми находится перинуклеарное пространство шириной от 20 до 60 нм. Оно заполнено энхилемой - сывороткообразной жидкостью с низкой электронной плотностью.
Итак, ядерная мембрана представляет собой полый мешок, отделяющий содержимое ядра от цитоплазмы, и состоит из двух слоев: внешний слой ограничивает перинуклеарное пространство снаружи, т. е. со стороны цитоплазмы, внутренний - изнутри, т. е. со стороны ядра. Из всех внутриклеточных мембранных компонентов подобным строением мембран обладают ядро, митохондрии и пластиды.
Морфологическое строение каждого слоя такое же, как и внутренних мембран цитоплазмы. Отличительная особенность ядерной оболочки - наличие в ней пор - округлых перфораций, образующихся в местах слияния внешней и внутренней ядерных мембран. Размеры пор довольно стабильны (30-100 нм в диаметре), в то же время их число изменчиво и зависит от функциональной активности клетки: чем активнее идут в ней синтетические процессы, тем больше пор приходится на единицу поверхности клеточного ядра.
Обнаружено, что количество пор увеличивается в период реконструкции и роста ядра, а также при репликации ДНК. Одно из крупнейших открытий, сделанных с помощью электронной микроскопии, - обнаружение тесной взаимосвязи между ядерной оболочкой и эндоплазматической сетью. Поскольку ядерная оболочка и тяжи эндоплазматической сети во многих местах сообщаются между собой, перинуклеарное пространство должно содержать ту же сывороткообразную жидкость, что и полости между мембранами эндоплазматической сети.
При оценке функциональной роли ядерной оболочки большое значение приобретает вопрос о ее проницаемости, обусловливающей обменные процессы между ядром и цитоплазмой в связи с передачей наследственной информации. Для правильного понимания ядерно-цитоплазматических взаимодействий важно знать, насколько ядерная оболочка проницаема для белков и других метаболитов. Опыты показывают, что ядерная оболочка легко проницаема для относительно крупных молекул. Так, рибонуклеаза - фермент, гидролизующий рибонуклеиновую кислоту без выделения свободной фосфорной кислоты, - имеет молекулярную массу около 13000 и очень быстро проникает в ядро.
Даже в корешках, фиксированных видоизмененным методом замораживания, можно наблюдать, как окрашивание ядрышек подавляется во всех клетках уже через 1 ч после обработки рибонуклеазой.
1.5. КАРИОПЛАЗМА
Кариоплазма (ядерный сок, нуклеоплазма) - основная внутренняя среда ядра, она занимает все пространство между ядрышком, хроматином, мембранами, всевозможными включениями и другими структурами. Кариоплазма под электронным микроскопом имеет вид гомогенной или мелкозернистой массы с низкой электронной плотностью. В ней во взвешенном состоянии находятся рибосомы, микротельца, глобулины и различные продукты метаболизма.
Вязкость ядерного сока примерно такая же, как вязкость основного вещества цитоплазмы. Кислотность ядерного сока, определенная путем микроинъекции индикаторов в ядро, оказалась несколько выше, чем у цитоплазмы.
Кроме того, в ядерном соке содержатся ферменты, участвующие в синтезе нуклеиновых кислот в ядре и рибосомы. Ядерный сок не окрашивается основными красителями, поэтому его называют ахроматиновым веществом, или кариолимфой, в отличие от участков, способных окрашиваться, - хроматина.
1.6. ХРОМАТИН
Термин «хромосома» используется по отношению к молекуле нуклеиновой кислоты, которая представляет собой хранилище генетической информации вируса, прокариота или эукариотической клетки. Однако первоначально слово «хромосома» (т. е. «окрашенное тело») использовалось в другом смысле, - для обозначения густо окрашенных образований в эукариотических ядрах, которые можно было наблюдать в световой микроскоп после обработки клеток красителем.
Эукариотические хромосомы, в изначальном смысле этого слова, выглядят как резко очерченные структуры только непосредственно до и во время митоза - процесса деления ядра в соматических клетках. В покоящихся, неделящихся эукариотических клетках хромосомный материал, называемый хроматином, выглядит нечетко и как бы беспорядочно распределен по всему ядру. Однако, когда клетка готовится к делению, хроматин уплотняется и собирается в свойственное данному виду число хорошо различимых хромосом.
Хроматин был выделен из ядер и проанализирован. Он состоит из очень тонких волокон, которые содержат 60 % белка, 35 % ДНК и, вероятно, 5 % РНК. Хроматиновые волокна в хромосоме свернуты и образуют множество узелков и петель. ДНК в хроматине очень прочно связана с белками, называемыми гистонами, функция которых состоит в упаковке и упорядочении ДНК в структурные единицы - нуклеосомы. В хроматине содержится также ряд негистоновых белков. В отличие от эукариотических, бактериальные хромосомы не содержат гистонов; в их состав входит лишь небольшое количество белков, способствующих образованию петель и конденсации (уплотнению) ДНК.
Глава 2. КЛЕТОЧНОЕ ЯДРО - ЦЕНТР УПРАВЛЕНИЯ ЖИЗНЕДЕЯТЕЛЬНОСТЬЮ КЛЕТКИ
2.1. ЯДРО - НЕЗАМЕНИМЫЙ КОМПОНЕНТ КЛЕТКИ
Еще в конце прошлого века было доказано, что лишенные ядра фрагменты, отрезанные от амебы или инфузории, через более или менее короткое время погибают. Более детальные опыты показали, что энуклеированные амебы живут, но вскоре после операции перестают питаться, двигаться и через несколько дней (до одной недели) погибают. Если пересадить ядро в ранее энуклеированную клетку, то процессы нормальной жизнедеятельности восстанавливаются и через некоторое время амеба начинает делиться.
Яйцеклетки морского ежа, лишенные ядра, при стимуляции к партеногенетическому развитию делятся, но тоже в конце концов погибают. Особенно интересные опыты были проведены на крупной одноклеточной водоросли ацетабулярии. После удаления ядра водоросль не только живет, но и в течение определенного периода может восстанавливать безъядерные участки. Следовательно, при отсутствии ядра прежде всего нарушается способность к размножению, и, хотя жизнеспособность на какое-то время сохраняется, в конце концов такая клетка неизбежно погибает.
содержание ядерного и лишенного ядра фрагмента в среде с радиоактивным предшественником РНК - 3Н-уридином показало, что синтез РНК в безъядерном фрагменте отсутствует. Белковый же синтез продолжается некоторое время за счет информационных РНК и рибосом, сформированных ранее, до удаления ядра. Пожалуй, наиболее яркую иллюстрацию роли ядра дают безъядерные эритроциты млекопитающих. Это эксперимент, поставленный самой природой.
Созревая, эритроциты накапливают гемоглобин, затем выбрасывают ядро и в таком состоянии живут и функционируют в течение 120 дней. Они не способны размножаться и в конце концов погибают. Однако клетки, только что выбросившие ядро, так называемые ретикулоциты, еще продолжают синтез белка, но уже не синтезируют РНК. Следовательно, удаление ядра влечет за собой прекращение поступления в цитоплазму новых РНК, которые синтезируются на молекулах ДНК, локализованных в хромосомах ядра. Однако это не мешает уже существующей в цитоплазме информационной РНК продолжать синтезировать белок, что и наблюдается в ретикулоцитах. Затем, когда РНК распадается, синтез белка прекращается, но эритроцит еще продолжает жить долгое время, выполняя свою функцию, которая не связана с интенсивным расходованием белка.
Лишенные ядра яйцеклетки морского ежа продолжают жить и могут делиться благодаря тому, что во время овогенеза запасли значительное количество РНК, которая и продолжает функционировать. Информационная РНК у бактерий функционирует минуты, но в ряде специализированных клеток млекопитающих она сохраняется сутки и больше.
Несколько особняком стоят данные, полученные на ацетобулярии. Оказалось, что морфогенез удаленной части определяется ядром, но жизнь кусочка обеспечивается ДНК, которую содержат хлоропласты. На этой ДНК синтезируется информационная РНК, которая, в свою очередь, обеспечивает синтез белка.
2.2. ФУНКЦИОНАЛЬНАЯ СТРУКТУРА ЯДРА
В изучении структурно-биохимической организации ядерного аппарата различных клеток большую роль играют сравнительно-цитологические исследования, в которых применяются как традиционный эволюционно-исторический подход, так и широкие сравнительно-цитологические сопоставления организации ядерного аппарата различных разновидностей клеток. Эволюционно-историческое направление в этих исследованиях имеет особое значение, поскольку ядерный аппарат представляет собой наиболее консервативную клеточную структуру - структуру, ответственную за хранение и передачу генетической информации.
Широкое сравнительно-цитологическое изучение ядерного аппарата у тех клеток, которые как бы резко уклоняются от обычного (типичного) уровня организации (ооциты, сперматозоиды, ядерные эритроциты, инфузории и т. д.), и использование данных, полученных с помощью молекулярно-биологических и цитологических методов в специальных науках, занимающихся клеточным уровнем организации (частная цитология, протозоология и т. д.), позволили выявить массу интересных особенностей организации ядерного аппарата, имеющих общецитологическое значение.
В составе ядерного аппарата эукариотных клеток можно выделить ряд субсистем, центральное место среди которых занимает совокупность интерфазных хромосом, или ДНК ядра. В них сосредоточена вся ДНК ядра, находящаяся в весьма сложных взаимоотношениях с белками хроматина, которые, в свою очередь, подразделяются на структурные, функциональные и регуляторные белки.
Второй и весьма важной субсистемой ядерного аппарата является ядерный матрикс, представляющий собой систему фибриллярных белков, выполняющих как структурную (скелетную) функцию в топографической организации всех ядерных компонентов, так и регуляторную функцию в организации процессов репликации, транскрипции, в созревании (процессинге) и перемещении продуктов транскрипции внутри ядра и за его пределы. По-видимому, белковый матрикс имеет двоякую природу: какие-то одни его компоненты обеспечивают в основном скелетную функцию, другие - регуляторную и транспортную.
Вместе с определенными участками ДНК хроматина белки ядерного матрикса (функционального и структурного) образуют основу ядрышка. Белки структурного матрикса принимают участие и в формировании поверхностного аппарата ядра. Поверхностный аппарат ядра занимает и в структурном, и в функциональном отношениях промежуточное положение между метаболическим аппаратом цитоплазмы и ядром. Мембраны и цистерны ядерной оболочки являются по сути дела специализированной частью общей мембранной системы цитоплазмы.
Специфическими структурами поверхностного аппарата ядра, играющими важную роль в реализации его основной функции - обеспечении взаимодействия ядра и цитоплазмы выступают поровые комплексы и субмембранная плотная пластинка, которые образуются с помощью белков ядерного матрикса. Наконец, последней субсистемой ядерного аппарата является кариоплазма. Это аналогичная гиалоплазме внешне бесструктурная фаза ядерного аппарата, которая создает специфическое для ядерных структур микроокружение, что обеспечивает возможность их нормального функционирования.
Кариоплазма находится в постоянном взаимодействии с гиалоплазмой через систему поровых комплексов и мембран ядерной оболочки.
2.3. РОЛЬ ЯДЕРНЫХ СТРУКТУР В ЖИЗНЕДЕЯТЕЛЬНОСТИ КЛЕТКИ
Основные процессы, связанные с синтезом белка, в принципе одинаковы у всех форм живого, указывают на особое значение клеточного ядра. Ядро осуществляет две группы общих функций: одну, направленную на собственно хранение генетической информации, другую - на ее реализацию, на обеспечение синтеза белка. Иными словами, первую группу составляют процессы поддержания наследственной информации в виде неизменной структуры ДНК. Эти процессы обусловлены наличием так называемых репарационных ферментов, ликвидирующих спонтанные повреждения молекул ДНК (разрыв одной из цепей ДНК, часть радиационных повреждений), что сохраняет строение молекул ДНК практически неизменными в ряду поколений клеток или организмов.
Далее в ядре происходит воспроизведение, или редупликация, молекул ДНК, что дает возможность двум клеткам получить совершенно одинаковые и в качественном, и в количественном смысле объемы генетической информации. В ядрах происходят процессы изменения и рекомбинации генетического материала, что наблюдается во время мейоза (кроссинговер). Наконец, ядра непосредственно участвуют в процессах распределения молекул ДНК при делении клеток.
Другой группой клеточных процессов, обеспечивающихся активностью ядра, является создание собственно аппарата белкового синтеза. Это не только синтез, транскрипция на молекулах ДНК разных информационных РНК, но транскрипция всех видов трансферных РНК и рибосомных РНК. В ядре эукариотов происходит также образование субъединиц рибосом путем комплексирования синтезированных в ядрышке рибосомных РНК с рибосомными белками, которые синтезируются в цитоплазме и переносятся в ядро. Таким образом, ядро представляет собой не только вместилище генетического материала, но и место, где этот материал функционирует и воспроизводится. Поэтому выпадение или нарушение любой из перечисленных выше функций гибельно для клетки в целом.
Так, нарушение репарационных процессов будет приводить к изменению первичной структуры ДНК и автоматически - к изменению структуры белков, что непременно скажется на их специфической активности, которая может просто исчезнуть или измениться так, что не сможет обеспечивать клеточные функции, в результате чего клетка погибает. Нарушения редупликации ДНК приведут к остановке размножения клеток или к появлению клеток с неполноценным набором генетической информации, что тоже гибельно для них. К такому же результату приведет нарушение процессов распределения генетического материала (молекул ДНК) при делении клеток.
Выпадение в результате поражения ядра или в случаях нарушений каких-либо регуляторных процессов синтеза любой формы РНК автоматически приведет к остановке синтеза белка в клетке или к грубым его нарушениям. Все это указывает на ведущее значение ядерных структур в процессах, связанных с синтезом нуклеиновых кислот и белков, главных функционеров в жизнедеятельности клетки.
Ядро осуществляет сложную координацию и регуляцию процессов синтеза РНК. Как указывалось, все три типа РНК образуются на ДНК. Радиографическими методами показано, что синтез РНК начинается в ядре (хроматине и ядрышке), и уже синтезированная РНК перемещается в цитоплазму. Таким образом мы видим, что ядро программирует синтез белка, который осуществляется в цитоплазме. Однако само ядро также испытывает влияние цитоплазмы, т. к. синтезируемые в ней ферменты поступают в ядро и необходимы для его нормального функционирования. Например, в цитоплазме синтезируется ДНК-полимераза, без которой не может происходить авторепродукция молекул ДНК. Поэтому следует говорить о взаимном влиянии ядра и цитоплазмы, при котором главенствующая роль все же принадлежит ядру как хранителю наследственной информации, которая передается при делении от одной клетки к другой.
2.4. ВЕДУЩЕЕ ЗНАЧЕНИЕ ДНК
Основное биологическое значение ядерного аппарата определяется его главным компонентом - гигантскими молекулами ДНК, способными к репликации и транскрипции. Эти два свойства ДНК и лежат в основе двух важнейших функций ядерного аппарата любой клетки:
а) удвоения наследственной информации и передачи ее в ряду клеточных поколений;
б) регулируемой транскрипции участков молекул ДНК и транспорта синтезируемых РНК в цитоплазму клеток.
По характеру организации ядерного аппарата все клетки делятся на три группы: прокариотные, мезокариотные и эукариотные.
Клеткам прокариот свойственны отсутствие ядерной оболочки, укладка ДНК без участия гистонов, унирепликонный тип репликации ДНК, моноцистронный принцип организации транскрипции и ее регуляция преимущественно по принципу положительной и отрицательной обратной связи.
Клетки эукариот, напротив, отличаются наличием ядерной оболочки, точнее говоря, даже сложного поверхностного аппарата ядра и мультирепликонным типом репликации молекул ДНК, образующих набор хромосом. Упаковка этих молекул происходит с помощью комплекса белков. Характер упаковки подвергается циклическим изменениям, связанным с прохождением клетками закономерных фаз цикла репродукции. Процессы транскрипции ДНК и ее регуляции у эукариот значительно отличаются от таковых у прокариот.
Мезокариотные клетки по организации ядерного аппарата занимают как бы промежуточное положение между эукариотными и прокариотными клетками. У мезокариот, как и у эукариот, имеется хорошо развитый поверхностный аппарат ядра. Укладка в хромосомы молекул ДНК существенно отличается от организации ДНП в эукариотных клетках. Механизмы репликации и транскрипции ДНК у мезокариот выяснены слабо. Таким образом в клеточном ядре протекают важнейшие процессы, связанные с наследственным статусом организма, - peпликация (биосинтез ДНК) и транскрипция.
Кроме того, ядро является источником отдельных белков и ферментов, необходимых для жизнедеятельности дифференцированных тканей. Одновременно с потоком информации в клетку для обеспечения синтеза белков осуществляется обратная связь: цитоплазма - ядро, т. е. ядро функционирует в тесном взаимодействии с другими частями клетки, объединяя процессы ядерно-цитоплазматического транспорта и регуляторного взаимодействия с цитоплазмой клетки.
Ядро клетки - это одна из основных составных частей всех растительных и животных клеток, неразрывно связанная с обменом, передачей наследственной информации и др.
Форма ядра клетки варьирует в зависимости от типа клетки. Имеются овальные, шаровидные и неправильной формы - подковообразные или многолопастные ядро клетки (у лейкоцитов), четковидные ядра клетки (у некоторых инфузорий), разветвленные ядра клетки (в железистых клетках насекомых) и др. Величина ядра клетки различна, но обычно связана с объемом цитоплазмы. Нарушение этого соотношения в процессе роста клетки приводит к клеточному делению. Количество ядер клетки также неодинаково - большинство клеток имеет одно ядро, хотя встречаются двуядерные и многоядерные клетки (например, некоторые клетки печени и костного мозга). Положение ядра в клетке является характерным для клеток каждого типа. В зародышевых клетках ядро обычно находится в центре клетки, но может смещаться по мере развития клетки и образования в цитоплазме специализированных участков или отложения в ней резервных веществ.
В ядре клетки различают основные структуры: 1) ядерную оболочку (ядерную мембрану), через поры которой осуществляется обмен между ядром клетки и цитоплазмой [имеются данные, указывающие на то, что ядерная мембрана (состоящая из двух слоев) без перерыва переходит в мембраны эндоплазматической сети (см. ) и комплекса Гольджи]; 2) ядерный сок, или кариоплазму,- полужидкую, слабо окрашиваемую плазматическую массу, заполняющую все ядра клетки и содержащую в себе остальные компоненты ядра; 3) (см.), которые в неделящемся ядре видны только с помощью специальных методов микроскопии (на окрашенном срезе неделящейся клетки хромосомы обычно имеют вид неправильной сети из темных тяжей и зернышек, в совокупности называемых ); 4) одно или несколько сферических телец - ядрышек, являющихся специализированной частью ядра клетки и связанных с синтезом рибонуклеиновой кислоты и белков.
Ядро клетки обладает сложной химической организацией, в которой важнейшую роль играют нуклеопротеиды - продукт соединения с белками. В жизни клетки имеются два основных периода: интерфазный, или метаболический, и митотический, или период деления. Оба периода характеризуются главным образом изменениями в строении ядра клетки. В интерфазе ядро клетки находится в покоящемся состоянии и участвует в синтезе белков, регуляции формообразования, процессах секреции и других жизненных отправлениях клетки. В период деления в ядре клетки происходят изменения, приводящие к перераспределению хромосом и образованию дочерних ядер клетки; наследственная информация передается, таким образом, через ядерные структуры новому поколению клеток.
Ядра клетки размножаются только делением, при этом в большинстве случаев делятся и сами клетки. Обычно различают: прямое деление ядра клетки путем перешнуровки - амитоз и самый распространенный способ деления ядер клетки- типичное непрямое деление, или митоз (см.).
Действие ионизирующей радиации и некоторых других факторов способно изменять заключенную в ядре клетки генетическую информацию, приводя к различным изменениям ядерного аппарата, что иногда может приводить к гибели самих клеток или служить причиной наследственных аномалий у потомства (см. Наследственность), Поэтому изучение структуры и функций ядра клетки, особенно связей между хромосомными соотношениями и наследованием признаков, которыми занимается цитогенетика, имеет существенное практическое значение для медицины (см. ).
См. также Клетка.
Ядро клетки - важнейшая составная часть всех растительных и животных клеток.
Клетка, лишенная ядра или с поврежденным ядром, не способна нормально выполнять свои функции. Ядро клетки, точнее, организованная в его хромосомах (см.) дезоксирибонуклеиновая кислота (ДНК),- носитель наследственной информации, определяющей все особенности клетки, тканей и целого организма, его онтогенез и свойственные организму нормы реагирования на воздействия среды. Заключенная в ядре наследственная информация закодирована в составляющих хромосомы молекулах ДНК последовательностью четырех азотистых оснований: аденина, тимина, гуанина и цитозина. Эта последовательность является матрицей, определяющей структуру синтезируемых в клетке белков.
Даже самые незначительные нарушения структуры ядра клетки ведут к необратимым изменениям свойств клетки или к ее гибели. Опасность ионизирующих излучений и многих химических веществ для наследственности (см.) и для нормального развития плода имеет в своей основе повреждения ядер в половых клетках взрослого организма или в соматических клетках развивающегося эмбриона. В основе преобразования нормальной клетки в злокачественную также лежат определенные нарушения структуры ядра клетки.
Размеры и форма ядра клетки и соотношение его объема и объема всей клетки характерны для различных тканей. Одним из главных признаков, отличающих элементы белой и красной крови, являются форма и размер их ядер. Ядра лейкоцитов могут быть неправильной формы: изогнуто-колбасовидной, лапчатой или четковидной; в последнем случае каждый участок ядра соединен с соседним тонкой перемычкой. В зрелых мужских половых клетках (сперматозоидах) ядро клетки составляет подавляющую часть всего объема клетки.
Зрелые эритроциты (см.) человека и млекопитающих не имеют ядра, так как они теряют его в процессе дифференцировки. Они имеют ограниченный срок жизни и не способны размножаться. В клетках бактерий и сине-зеленых водорослей отсутствует резко очерченное ядро. Однако в них содержатся все характерные для ядра клетки химические вещества, распределяющиеся при делении по дочерним клеткам с такой же правильностью, как и в клетках высших многоклеточных организмов. У вирусов и фагов ядро представлено единственной молекулой ДНК.
При рассмотрении покоящейся (неделящейся) клетки в световом микроскопе ядро клетки может иметь вид бесструктурного пузырька с одним или несколькими ядрышками. Ядро клетки хорошо красится специальными ядерными красками (гематоксилин, метиленовый синий, сафранин и др.), которые обычно используют в лабораторной практике. При помощи фазово-контрастного устройства ядро клетки можно исследовать и прижизненно. В последние годы для изучения процессов, протекающих в ядре клетки, широко используют микрокинематографию, меченые атомы С14 и Н3 (ауторадиография) и микроспектрофотометрию. Последний метод особенно успешно применяют для изучения количественных изменений ДНК в ядре в процессе жизненного цикла клетки. Электронный микроскоп позволяет выявить детали тонкой структуры ядра покоящейся клетки, необнаруживаемые в оптическом микроскопе (рис. 1).
Рис. 1. Современная схема строения клетки, основанная на наблюдениях в электронном микроскопе: 1 - цитоплазма; 2 - аппарат Гольджи; 3 - центросомы; 4 - эндоплазматический ретикулум; 5 - митохондрии; 6 - оболочка клетки; 7 - оболочка ядра; 8 - ядрышко; 9 - ядро.
При делении клеток - кариокинезе или митозе (см.) - ядро клетки претерпевает ряд сложных преобразований (рис. 2), во время которых становятся отчетливо видимыми его хромосомы. Перед делением клетки каждая хромосома ядра синтезирует из веществ, присутствующих в ядерном соке, себе подобную, после чего материнская и дочерняя хромосомы расходятся к противоположным полюсам делящейся клетки. В результате каждая дочерняя клетка получает такой же хромосомный набор, какой был у материнской клетки, а вместе с ним и заключенную в нем наследственную информацию. Митоз обеспечивает идеально правильное разделение всех хромосом ядра на две равнозначные части.
Митоз и мейоз (см.) являются важнейшими механизмами, обеспечивающими закономерности явлений наследственности. У некоторых простейших организмов, а также в патологических случаях в клетках млекопитающих и человека ядра клетки делятся путем простой перетяжки, или амитоза. В последние годы показано, что и при амитозе происходят процессы, обеспечивающие разделение ядра клетки на две равнозначные части.
Набор хромосом в ядре клетки особи называют кариотипом (см.). Кариотип во всех клетках данной особи, как правило, одинаков. Многие врожденные аномалии и уродства (синдромы Дауна, Клайнфелтера, Тернера-Шерешевского и др.) обусловлены различными нарушениями кариотипа, возникшими либо на ранних стадиях эмбриогенеза, либо при созревании половой клетки, из которой возникла аномальная особь. Аномалии развития, связанные с видимыми нарушениями хромосомных структур ядра клетки, называют хромосомными болезнями (см. Наследственные болезни). Различные повреждения хромосом могут быть вызваны действием физических или химических мутагенов (рис. 3). В настоящее время методы, позволяющие быстро и точно устанавливать кариотип человека, используют для ранней диагностики хромосомных болезней и для уточнения этиологии некоторых заболеваний.

Рис. 2. Стадии митоза в клетках культуры ткани человека (перевиваемый штамм НЕр-2): 1 - ранняя профаза; 2 - поздняя профаза (исчезновение ядерной оболочки); 3 - метафаза (стадия материнской звезды), вид сверху; 4 - метафаза, вид сбоку; 5 - анафаза, начало расхождения хромосом; 6 - анафаза, хромосомы разошлись; 7 - телофаза, стадия дочерних клубков; 8 - телофаза и разделение клеточного тела.

Рис. 3. Повреждения хромосом, вызываемые ионизирующей радиацией и химическими мутагенами: 1 - нормальная телофаза; 2-4 - телофазы с мостами и фрагментами в эмбриональных фибробластах человека, облученных рентгеновыми лучами в дозе 10 р; 5 и 6 - то же в кроветворных клетках морской свинки; 7 - хромосомный мост в эпителии роговицы мыши, облученной дозой в 25 р; 8 - фрагментация хромосом в эмбриональных фибробластах человека в результате воздействия нитрозоэтилмочевиной.
Важный органоид ядра клетки - ядрышко - является продуктом жизнедеятельности хромосом. Оно продуцирует рибонуклеиновую кислоту (РНК), являющуюся обязательным промежуточным звеном в синтезе белка, вырабатываемого каждой клеткой.
Ядро клетки отделено от окружающей цитоплазмы (см.) оболочкой, толщина которой 60-70 Å.
Через поры в оболочке вещества, синтезируемые в ядре, поступают в цитоплазму. Пространство между оболочкой ядра и всеми его органоидами заполнено кариоплазмой, состоящей из основных и кислых белков, ферментов, нуклеотидов, неорганических солей и других низкомолекулярных соединений, необходимых для синтеза дочерних хромосом при делении ядра клетки.
Ядро, его строение и биологическая роль.
Ядро состоит из 1)поверхн аппарата ядра (в нем выдлел: 2 мембраны, перинуклеарн пространств, поровые комплексы, ламину.) 2) кариоплазмы (нуклеоплазмы) 3) хроматина (в нём эухроматин и гетерохроматин) 4) ядрышка (грануляр и фибриляр компонент.)
Ядро – это структура клетки которая выполняет функцию хранения и передачи инф, а так же регулирует все жизненные процессы клетки. Ядро несёт в себе генетическую (наследственную) инф в виде ДНК. Ядра обычно имеют шаровидную или яйцевидную форму. Я. окружено ядерн оболочкой. Ядерная оболочка пронизана ядерными порами. Через них ядро обменивается веществами с цитоплазмой(внутр средой клетки). Наружная мембрана переходит в эндоплпзматич ретикулум и может быть усеяна рибосомами. Отношение размеров ядра и клетки зависит от функциональной активности клетки. Большинство клеток одноядерные. Двуядерными могут быть кардиомиоциты. Всегда двуядерны инфузории. В них характерен ядерный дуализм.(то есть ядра различ по строению и финкциям). Малое ядро (генеративное) – диплойдное. Оно обеспечивает только половой процесс у инфузорий. Большое (вегетативное) ядро полиплойдное. Оно регулирует все остальные жизненные процессы. Многоядерными бывают клетки некоторых простейших и клетки скелетной мускулатуры.
П.А.Я. или кариотека ) имеет микроскопическую толщину и поэтому виден в световой микроскоп. Поверхностный аппарат ядра включает:
а)ядерную оболочку, или кариолемму;. б)паровые комплексы; в)периферическую плотную пластинку (ППП), или ламину.
(1) Ядерная оболочка (кариолемма). состоит из 2 мембран - наружной и внутренней, разделённых перинукляеарным пространством. Обе мембраны имеют такое же жидкосто-мозаичное строение, как и плазматическая мембрана, и различаются по набору белков. Среди этих белков имеются ферменты, переносчики и рецепторы. Наружная ядерная мембрана является продолжением мембран грЭПС и может быть усеяна рибосомами, на которых идёт синтез белка. Со стороны цитоплазмы наружная мембрана окружена сетью промежуточных (ви-ментиновых) фипаментов. Между наружной и внутренней мембранами находится перинуклеарное пространство -полость шириной 15-40 нм, содержимое которого сообщается с полостями каналов ЭПС. По составу перинуклеарное пространство близко к гиалоплазме и может содержать синтезированные рибосомами белки. Главная функция кариолеммы - изоляция гиалоплазмы от кариоплазмы. Специальные белки ядерных мембран, расположенные в области ядерных пор, осуществляют транспортную функцию. Ядерная оболочка пронизана ядерными порами, через которые осуществляется связь кариоплазмы и гиалоплазмы. Для регуляции такой связи в порах находятся (2) поровые комплексы. Они занимают 3-35% поверхности ядерной оболочки. Число ядерных пор с поровыми комплексами является изменчивой величиной и зависит от активности ядра. В области ядерных пор наружная и внутренняя ядерные мембраны сливаются. Совокупность структур, связанных с ядерной порой, называется комплексом ядерной поры. Типичный поровый комплекс представляет собой сложную белковую структуру - содержит более 1000 молекул белка. В центре поры расположена центральная белковая глобула (гранула), от которой по радиусу отходят тонкие фибриллы к периферическим белковым глобулам, образуя диафрагму поры. По периферии ядерной поры находятся две параллельные кольцевые структуры диаметром 80-120 нм (по одному с каждой поверхности кариолеммы), каждое из которых образовано 8 белковыми гранулами (глобулами).
Белковые глобулы перового комплекса подразделяются на центральные и периферические . С помощью периферических глобул осуществляется транспорт макромолекул из ядра в гиалоплазму. (фиксируются в мембране специальным интегральным белком. От этих гранул к центру сходятся белковые фибриллы, формирующие перегородку - диафрагму поры)
В нем участвуют специальные белки периферических глобул - нуклеопорины. В периферических глобулах имеется особый белок - переносчик молекул т-РНК
Центральная глобула специализируется на транспорте и-РНК из ядра в гиалопдазму. В её составе имеются ферменты, участвующее в химической модификации иРНК - ее процессинге.
Гранулы поровых комплексов структурно связаны с белками ядерной ламины, которая участвует в их организации
Функции комплекса ядерной поры:
1. Обеспечение регуляции избирательного транспорта в-в между цитоплазмой и ядром.
2. Активный перенос в ядро белков
3. Перенос в цитоплазму субъединиц рибосом
(3) ППП или ламина
слой толщиной 80-300 нм. прилегает изнутри к внутренней ядерной мембране. Внутренняя ядерная мембрана гладкая, ее интегральные белки связаны с ламиной (периферической плотной пластинкой). Ламина состоит из специальных переплетенных белков-ламинов, образующих периферический кариоскелет. Белки-ламины относятся к классу промежуточных филаментов (скелетных фибрилл). У млекопитающих известно 4 вида этих белков - это ломимы А, В, В 2 и С. Эти белки поступают в ядро из цитоплазмы. Ламины разных видов взаимодействуют между сбой и образуют белковую сеть под внутренней мембраной ядерной оболочки. С помощью ламинов «В» ППП соединяется со спец интеграл белкомядерн оболочки. С ППП взаимодействуют и белки приферич голобул «внутр кольца» порового комплекса. К ламину «А» присоед теломерн участки хромосом.
Функции ламины: 1) поддерд форму ядра. (даже есл бое мембраны разруш, то ядро за счет ламины сохр свою форму и поровые комп-сы ост на своём месте.
2) служит компонентом кариоскелета
3) участв в сборке ядерн оболочки (формирование кариоллемы) при делен клетки.
4) в интерфазном ядре к ламине прикрепл хроматин. таким образом ламина обеспеч функцию фиксации хроматина в ядре (обеспеч упорядочн укладку хроматина, участвует в пространственной организации хроматина в интерфазном ядре). Ламин «А» взаимодейств с теломерными участками хромосом.
5) обеспеч структур организацию поровых комплексов.
импорт и экспор белков.
В ядро через ядерные поры поступают: синтезированные цитоплазматическими рибосомами белки-ферменты, которые участвуют в процессах репликации и репарации (восстановления повреждений в ДНК); белки-ферменты, участвующие в процессе транскрипции; белки-репрессоры, которые регулируют процесс транскрипции; белки-гистоны.(которые связаны с молекулой ДНК и образуют хроматин); белки, входящие в состав субъединиц рибосом: белки ядерного матрикса, образующие кариоскелет; нуклеотиды; ионы минеральных солей, в частности, ионы Са и Mg .
Из ядра в цитоплазму выходят и-РНК. т-РНК и субъединицы рибосом, которые представляют собой рибонуклеопротеидные частицы (р-РНК, связанные с белками).
5. Химический состав и структурная организация хроматина. уровни компактизации. хромосомы чел их строен и классификация.
В ядре клеток мелкие зернышки и глыбки материала, окрашиваются основными красителями.
Хроматин представляет собой дезоксирибонуклеопротеид (ДНП) и состоит из ДНК, соединённой с белка-ми-гистонами или негистоновыми белками. Гистоны и ДНК объединены в структуры, которые называются нуклеосомами. Хроматин соответствует хромосомам, которые в интерфазном ядре представлены длинными перекрученными нитями и неразличимы как индивидуальные структуры. Выраженность спирализации каждой из хромосом неодинакова по их длине. Реализацию генетической информации осуществляют деспирализованные участки хромосом.
классификация хроматина:
1) эухроматин (активный деспирализованный. на нем происход считывание инф (транскрипция). в ядре выявляется как более светлые участки ближе к центру ядра) Предполагается, что в нем сосредоточена та ДНК, которая в интерфазе генетически активна. Эухроматин соответствует сегментам хромосом, которые деспирализованы и открыты для транскрипции.
2) гетерохроматин (нерабочий спирализованный, конденсированный, более компактный В ядре выявляется в виде глыбок на периферии.) делится на: конститутивный (всегда неактивен, никогда не переходит в эухроматин) и Факультативный (при определён условиях или на определен стадиях иммунного цикла может переходить в эухроматин). располагается ближе к оболочке ядра, более компактный. Примером скопления факульт гетерохроматина является тельце Барра - инактивированная Х-хромосома у самок млекопитающих, которая в интерфазе плотно скручена и неактивна.
Таким образом, по морфологическим признакам ядра (по соотношению содержания эу- и гетерохроматина) можно оценить активность процессов транскрипции, а, следовательно, синтетической функции клетки.
Хроматин и хромосомы представляют собой дезоксирибонуклеопротеиды (ДНП), но хроматин - это раскрученное, а хромосомы - скрученное состояние. Хромосом в интерфазном ядре нет, хромосомы появляются при разрушении ядерной оболочки (во время деления).
Строение хромосом:
хромосомы - наиболее упакованное состояние хроматина.
В хромосомах различают первичную перетяжку (центромеру), разделяющую хромосому на два плеча. Первичная перетяжка - наименее спирализованная часть хромосомы, к ней во время деления клетки присоединяются нити веретена деления. На некоторых хромосомах есть глубокие вторичные перетяжки, отделяющие небольшие участки хромосом, называемые спутниками. В области вторичных перетяжек находятся гены, кодирующие информацию об р-РНК, поэтому вторичные перетяжки хромосом называются ядрышковыми организаторами.
В зависимости от места расположения центромеры различают три типа хромосом:
1) метацентрические (имеют плечи равной или почтиравной величины);
2) субметацентрические (имеют плечи неравной величины);
3) акроцентрические (имеют палочковидную форму с коротким, почти незаметным вторым плечом);
Концы плеч хромосом называются теломерами
Уровни компаюпизации хроматина:
1. Нуклеосомный - Два с половиной витка двойной спирали ДНК (в 146-200 пар нуклеотидов) наматываются снаружи на белковый кор, образуя нуклеосому. Каждый гистон представлен двумя молекулами. ДНК наматывается на кор снаружи, образуя два с половиной витка. Участок ДНК между нуклеосомами называется линкером и имеет протяжбенность 50-60 пар нуклеотидов. Толщина нуклеосомной нити составляет 8-11 нм.
2. Нуклеомерный. Нуклеосомная структура закручивается, образуя суперспираль. В её образовании принимает участие ещё один гистоновый белок HI, лежащий между нуклеосомами и связанный с линкером. К каждому линкеру присоединяется 1 молекула гистона HI. Молекулы HI в комплексе с линкерами взаимодействуют между собой и вызывают суперспирализацию нуклеосомной фибриллы.
В результате образуется хроматиновая фибрилла, толщина которой составляет 30 нм (ДНК компактизирована в 40 раз). Суперспирализация происходит двумя способами. 1) нуклеосомная фибрилла может образовывать спираль второго порядка, которая имеет форму соленоида; 2) 8-10 нуклеосом образуют крупную компактную структуру - нуклеомеру. Этот уровень не допускает синтеза РНК с нуклеомерной ДНК (транскрипция не происходит).
3. Хромомерный (петельная структура). Хроматиновая фибрилла образует петли, которые сцепляются между собой с помощью особых негистоновых белков, либо петельные центры - хромомеры. Толщина 300 нм.
4. Хромонемный - образуется в результате сближения хромомеров по длине. Хромонема содержит одну гигантскую молекулу ДНК в комплексе с белками, т.е. фибриллу дезокси-рибонуклеопротеина - ДНП (400 нм).
5. Хроматидный - хромонема складывается несколько раз, образуя тело хроматиды (700 нм). После репликации ДНК хромосома содержит 2 хроматиды.
6. Хромосомный (1400 нм). Состоит из двух хроматид. Хроматиды соединены центромерой. При делении клетки хроматиды расходятся, попадая в разные дочерние клетки.
хромосомы человека
Кариоти́п - совокупность признаков (число, размеры, форма и т.д.) полного набора хромосом, присущий клеткам данного биологического вида (видовой кариотип ), данного организма (индивидуальный кариотип ) или линии (клона) клеток.
Для процедуры определения кариотипа могут быть использованы любые популяции делящихся клеток, для определения человеческого кариотипа используется либо одноядерные лейкоциты, извлечённые из пробы крови, деление которых провоцируется добавлением митогенов, либо культуры клеток, интенсивно делящихся в норме (фибробласты кожи, клетки костного мозга).
кариотип – диплойдный набор хромосом, свойтвенный соматическим клеткам организмов данного вида, являющийся видоспецифическим признаком и характеризующийся определённым числом и строением хромосом.
Хромосомный набор большинства клеток - диплоидный (2п) - это значит, что каждая хромосома имеет пару, т.е. гомологичную хромосому. Обычно диплоидный (2п) набор хромосом образуется в момент оплодотворения (одна из пары хромосом от отца, другая от матери). Некоторые клетки триплоидны (Зп), например клетки эндосперма.
Изменение числа хромосом в кариотипе человека может привести к различным заболеваниям. Наиболее частым хромосомным заболеванием у человека является синдром Дауна , обусловленный трисомией (к паре нормальных хромосом прибавляется еще одна такая же, лишняя) по 21-й хромосоме. Встречается этот синдром с частотой 1-2 на 1000.
Известны трисомии по 13-й хромосоме - Синдром Патау
, а также по 18-й хромосоме - синдром Эдвардса
, при которых жизнеспособность новорожденных резко снижена. Они гибнут в первые месяцы жизни из-за множественных пороков развития.
Достаточно часто у человека встречается изменение числа половых хромосом. Среди них известна моносомия Х (из пары хромосом присутствует только одна (Х0)) - это синдром Шерешевского-Тернера
. Реже встречается трисомия Х и синдром Клайнфельтера
(ХХУ, ХХХУ, ХУУ и т.д.)
6. Гиалоплазма. Органеллы, их классификация. Биологические мембраны.
гиалоплазма - часть цитоплазмы животных и растительных клеток, не содержащая структур, различимых в световом микроскопе.
Гиалоплазма (hyaloplasma; от греч. hyalinos - прозрачный) составляет примерно 53-55 % от общего объема цитоплазмы (cytoplasma), образуя гомогенную массу сложного состава. В гиалоплазме присутствуют белки, полисахариды, нуклеиновые кислоты, ферменты. При участии рибосом в гиалоплазме синтезируются белки, происходят различные реакции промежуточного обмена. В гиалоплазме располагаются также органеллы, включения и клеточное ядро.
Основная роль гиалоплазмы – объединение всех клеточных структур в отношении их химического взаимодействия и обеспечения транспортных биохимических процессов.
Органеллы (organellae) являются обязательными микроструктурами для всех клеток, выполняющими определенные жизненно важные функции. Различают мембранные и немембранные органеллы .
К мембранным органеллам , отграниченным от окружающей их гиалоплазмы мембранами, относятся эндоплазматическая сеть, комплекс Гольджи, лизосомы, пероксисомы, митохондрии.
Эндоплазматическая сеть представляет собой единую непрерывную структуру, образованную системой цистерн, трубочек и уплощенных мешочков. На электронных микрофотографиях различают зернистую (шероховатую, гранулярную) и незернистую (гладкую, агранулярную) эндоплазматическую сеть. Внешняя сторона зернистой сети покрыта рибосомами, незернистая лишена рибосом. Зернистая эндоплазматическая сеть синтезирует (на рибосомах) и транспортирует белки. Незернистая сеть синтезирует липиды и углеводы и участвует в их обмене (например, стероидные гормоны в корковом веществе надпочечников и клетках Лейдига (сустеноцитах) яичек; гликоген - в клетках печени). Одной из важнейших функций эндоплазматической сети является синтез мембранных белков и липидов для всех клеточных органелл.
комплекс Гольджи представляет собой совокупность мешочков, пузырьков, цистерн, трубочек, пластинок, ограниченных биологической мембраной. Элементы комплекса Гольджи соединены между собой узкими каналами. В структурах комплекса Гольджи происходят синтез и накопление полисахаридов, белково-углеводных комплексов, которые выводятся из клеток. Так образуются секреторные гранулы. Комплекс Гольджи имеется во всех клетках человека, кроме эритроцитов и роговых чешуек эпидермиса. В большинстве клеток комплекс Гольджи расположен вокруг или вблизи ядра, в экзокринных клетках - над ядром, в апикальной части клетки. Внутренняя выпуклая поверхность структур комплекса Гольджи обращена в сторону эндоплазматической сети, а внешняя, вогнутая, - к цитоплазме.
Мембраны комплекса Гольджи образованы зернистой эндоплазматической сетью и переносятся транспортными пузырьками. От внешней стороны комплекса Гольджи постоянно отпочковываются секреторные пузырьки, а мембраны его цистерн постоянно обновляются. Секреторные пузырьки поставляют мембранный материал для клеточной мембраны и гликокаликса. Таким образом обеспечивается обновление плазматической мембраны.
Лизосомы представляют собой пузырьки диаметром 0,2-0,5 мкм, содержащие около 50 видов различных гидролитических ферментов (протеазы, липазы, фосфолипазы, нуклеазы, гликозидазы, фосфатазы). Лизосомальные ферменты синтезируются на рибосомах зернистой эндоплазматической сети, откуда переносятся транспортными пузырьками в комплекс Гольджи. От пузырьков комплекса Гольджи отпочковываются первичные лизосомы. В лизосомах поддерживается кислая среда, ее рН колеблется от 3,5 до 5,0. Мембраны лизосом устойчивы к заключенным в них ферментам и предохраняют цитоплазму от их действия. Нарушение проницаемости лизосомальной мембраны приводит к активации ферментов и тяжелым повреждениям клетки вплоть до ее гибели.
Во вторичных (зрелых) лизосомах (фаголизосомах) происходит переваривание биополимеров до мономеров. Последние транспортируются через лизосомальную мембрану в гиалоплазму клетки. Непереваренные вещества остаются в лизосоме, в результате чего лизосома превращается в так называемое остаточное тельце высокой электронной плотности.
Митохондрии (mitochondrii), являющиеся «энергетическими станциями клетки», участвуют в процессах клеточного дыхания и преобразования энергии в формы, доступные для использования клеткой. Их основные функции - окисление органических веществ и синтез аденозинтрифосфорной кислоты (АТФ). Много крупных митохондрий в кардиомиоцитах, мышечных волокнах диафрагмы. Они расположены группами между миофибриллами, окружены гранулами гликогена и элементами незернистой эндоплазматической сети. Митохондрии являются органеллами с двойными мембранами (толщина каждой около 7 нм). Между наружной и внутренней митохондриальными мембранами расположено межмембранное пространство шириной 10-20 нм.
К немембранным органоидам относятся клеточный центр эукариотических клеток и рибосомы, имеющиеся в цитоплазме как эу- , так и прокариотических клеток.
Рибосома - это округлая рибонуклеопротеиновая частица диаметром 20-30 нм. Она состоит из малой и большой субъединиц, объединение которых происходит в присутствии матричной (информационной) РНК (мРНК). Одна молекула мРНК обычно объединяет несколько рибосом наподобие нитки бус. Такую структуру называют полисомой. Полисомы свободно располагаются в основном веществе цитоплазмы или прикреплены к мембранам шероховатой цитоплазматической сети. В обоих случаях они служат местом активного синтеза белка.
70S - рибосомы обнаруживаются у прокариот и в хлоропластах и митохондриях эукариот. 8OS-рибосомы, несколько более крупные, находятся в цитоплазме эукариот. В процессе синтеза белка рибосомы движутся вдоль мРНК. Процесс идет более эффективно, если вдоль мРНК движется не одна, а несколько рибосом. Такие цепи рибосом на мРНК называют полирибосомами, или полисомами.
МЕМБРАНЫ:
все мембраны образуют липопротеидные плёнки; имеют двойной слой липидов.
В составе мембран до 20% воды. липиды.
Мембраны состоят из липидов трех классов: фосфолипиды, гликолипиды и холестерол. Фосфолипиды и гликолипиды состоят из двух длинных гидрофобных углеводородных «хвостов», которые связаны с заряженной гидрофильной «головой». Холестерол придает мембране жесткость, занимая свободное пространство между гидрофобными хвостами липидов и не позволяя им изгибаться. Поэтому мембраны с малым содержанием холестерола более гибкие, а с большим - более жесткие и хрупкие.
Клеточные мембраны часто асимметричны, то есть слои отличаются по составу липидов, переход отдельной молекулы из одного слоя в другой (так называемый флип-флоп ) затруднен. Различается состав и ориентация мембранных белков.
Одна из важнейших функций биомембраны - барьерная. Например, мембрана пероксисом защищает цитоплазму от опасных для клетки пероксидов.
Еще одно важное свойство биомембраны - избирательная проницаемость.







