Концепция единства материальных структур и онтологической безмассовой волновой среды позволяет понять природу всех типов взаимодействия и системную организацию структуры нуклонов, ядер и атомов. Нейтроны играют ключевую роль в формировании и сохранении стабильности ядер, которая обеспечивается двумя бозоно-обменными связями между протонами и нейтронами. Альфа-частицы – главные «кирпичики» в структуре. Структуры ядер, близкие по форме к сферической, образованы в соответствии с периодами в периодической системе Д.И. Менделеева последовательным добавлением комплекса n-p-n, альфа-частицы и нейтронов. Причиной радиоактивного распада атомов является не оптимальная структура ядра: превышение числа протонов или нейтронов, асимметрия. Альфа-структура ядер объясняет причины и энергетический баланс всех типов радиоактивного распада.
структура нуклонов
альфа-частиц
«бозоно-обменные» силы
стабильность
радиоактивность
1. Вернадский В.И. Биосфера и ноосфера. – М.: Рольф. 2002. – 576 с.
2. Дмитриев И.В. Вращение по одной, двум или трём собственным внутренним осям – необходимое условие и форма существования частиц физического мира. – Самара: Самарское кн. изд-во, 2001. – 225 с.
3. Поляков В.И. Экзамен на «Homo sapiens» (От экологии и макроэкологии… к МИРУ). – Саранск: изд-во Мордовского университета, 2004. – 496 с.
4. Поляков В.И. ДУХ МИРА вместо хаоса и вакуума (Физическая структура Вселенной) // «Современные наукоёмкие технологии».- –2004. №4. – С.17-20.
5. Поляков В.И. Электрон = позитрон?! //Современные наукоёмкие технологии. – 2005. – №11. – С. 71-72.
6. Поляков В.И. Рождение материи //Фундаментальные исследования 2007. №12. – С.46-58.
7. Поляков В.И. Экзамен на «Homo sapiens – II». От концепций естествознания ХХ века – к естествопониманию. – Изд-во «Академия естествознания». – 2008. – 596 с.
8. Поляков В.И. Почему стабильны протоны и радиоактивны нейтроны? // «Радиоактивность и радиоактивные элементы в среде обитания человека»: IV Международная Конференция, Томск, 5-7 июня 2013. – Томск, 2013. – С. 415-419.
9. Поляков В.И. Основы естествопонимания структуры нуклонов, ядер, стабильности и радиоактивности атомов // Там же. – С. 419-423.
10. Поляков В.И. Структуры атомов – орбитально волновая модель// Успехи современного естествознания. – 2014. №3. – С.108-114.
12. Физические величины: Справочник // А.П. Бабичев, Н.А. Бабушкина, А.М. Братковскийи др.; Под ред. И.С. Григорьева, Е.З. Мелихова. – М.: Энергоатомиздат, 1991. – 1232 с.
Современная физика предлагает для описания структуры ядер капельную, оболочечную, обобщённую и другие модели. Связь нуклонов в ядрах объясняется энергией связи, обусловленной «особыми специфическими ядерными силами» . Свойства этих сил (притяжение, короткодействие, зарядовая независимость и т.д.) приняты как аксиома. Вопрос «почему так?» возникает почти к каждому тезису. «Принято (?), что эти силы одинаковы для нуклонов… (?). Для лёгких ядер удельная энергия связи круто возрастает, претерпевая целый ряд скачков (?), затем более медленно возрастает (?), а потом постепенно уменьшается» . «Наиболее устойчивым оказываются так называемые «магические ядра», у которых число протонов или нейтронов равно одному из магических чисел: 2, 8, 20, 28, 50, 82, 126…(?) Особенно стабильны дважды магические ядра: 2He2, 8O8, 20Ca20, 20Ca28, 82Pb126» (левый и правый индексы соответствуют числу протонов и нейтронов в ядре, соответственно). Почему существуют «магические» ядра, а магический изотоп 28Ni28 с максимальной удельной энергией связи 8,7 МэВ - короткоживущий
(Т1/2 = 6,1 сут.)? «Ядра характеризуются практически постоянной энергией связи и постоянной плотностью, не зависящей от числа нуклонов» (?!). Это означает, что энергия связи ничего не характеризует, также как и табличные значения дефекта массы (у 20Са20 меньше, чем 21Sc24, у 28Ni30 меньше, чем 27Co32 и 29Cu34 и т.д.) . Физика признаёт, что «сложный характер ядерных сил и трудности решения уравнений…не позволили до настоящего времени разработать единую последовательную теорию атомного ядра» . Наука ХХ века, построенная на постулатах теории относительности, отменила логику и причинно-следственную связь, а математические фантомы объявила реальностью. Не познав структуры ядер и атомов, учёные создали атомные бомбы и пытаются в коллайдерах имитировать Большой взрыв Вселенной…
«Революция в естествознании А. Эйнштейна» подменила уравнениями «пространственно-временного континиума» труды десятков выдающихся учёных (Гюйгенс, Гук, Юнг, Навье, Стокс, Герц, Фарадей, Максвелл, Лоренц, Томсон, Тесла и др.), которые развивали теории электромагнетизма и атомизма в среде «эфир». Следует вернуться на век назад…
Цель и метод работы. Выход из тупика науки возможен на основе понимания сущности среды «эфир». В.И. Вернадский писал: «Излучениями НЕ МАТЕРИАЛЬНОЙ среды охвачено всё доступное, всё мыслимое пространство… Кругом нас, в нас самих, всюду и везде, без перерыва, вечно сменяясь, совпадая и сталкиваясь, идут излучения разной длины волны - от волн, длина которых исчисляется десятимиллионными долями миллиметра, до длинных, измеряемых километрами…Всё пространство ими заполнено…» . Всё материальное формируется этой онтологической, не материальной, волновой средой и существует во взаимодействии с ней. «Эфир» - это не газ и не хаос вихрей, а «Действие, Упорядочивающие Хаос - ДУХ» . В среде ДУХ из единственной элементарной частицы - массона (электрон/позитрон) закономерно и системно организованы структуры от нуклонов, ядер и атомов до Вселенной .
В работе развита модель структуры ядер, которая объясняет их свойства, причины связи нуклонов в ядрах, особую стабильность и радиоактивность.
Структура и свойства нуклонов
Принятая в физике модель нуклонов построена из десятков гипотетических частиц со сказочным названием «кварк» и сказочными отличиями, среди которых: цвет, очарование, странность, прелесть . Эта модель слишком сложна, не имеет доказательств и не может объяснить даже массу частиц. Модель структуры нуклонов, объясняющая все их свойства, была разработана И.В. Дмитриевым (г. Самара) на основе экспериментально открытого им принципа максимума конфигурационной энтропии (равенство структурных элементов на поверхности и в объёме первочастиц) и тезиса о существовании частиц только при вращении «по одной, двум или трём собственным внутренним осям» . Нуклон сформирован из 6-и гексагональных структур π+(-)-мезонов, окружающих плюс-мюон μ+, а их структура построена подбором количества шаров: электронов и позитронов двух типов. Такая структура была обоснована на основе взаимодействия материальных частиц массонов и среды ДУХ в работе , а затем уточнена и доказана на основе построения структуры мезонов в соответствии с постоянной тонкой структуры
1/α = 2h(ε0/μ0)1/2/e2 = 137,036 . Над физическим смыслом этой константы ломали головы физики В. Паули, Р. Фейнман), а в среде ДУХ он очевиден: только на относительном расстоянии 1/α от заряда существует волновое взаимодействие материи и среды.
Расчётное число массонов (me) в структуре мюона должно быть 3/2α = 205,6 , а масса мюона 206,768 me . В его структуре из 207 массонов, центральный определяет заряд ±e и спин ±1/2, а 206 взаимно компенсируются. Пионы, как постулировано И. Дмитриевым, сформированы из «двухосных» электронов и позитронов (спин = 0, заряд +/-, масса me). В среде ДУХ бозоны с массой 2/3 me должны образовываться как первый этап формирования материи из квантов фонового излучения Вселенной в атмосфере Солнца . Таких частиц в плотной структуре должно быть 3/α = 411 частиц, а их масса должна составлять 3/α · 2/3 me = 274 me , что соответствует пи-мезонам (mπ = 273,210 me ). Их структура подобна мюонам: частица в центре определяет заряд ± 2/3e и спин 0, а 205 частицы взаимно уравновешены.
Структура протона из центрального мюона и 6 пионов, с учётом потери массы на обменную («ядерную») связь из 6 массонов (связь мюона с пионами) и 6 бозонов (связь между пионами, 4 me) объясняет его массу.
Мр = 6mp + mm - 10me = 6·273,210 me+ +206,768 me - 10me =1836,028 me.
Это значение с точностью 0,007 % соответствует массе протона Мр = 1836,153me . Заряд протона +e и спин ±1/2 определяются центральным массоном+ в центральном мюоне+. Модель протона объясняет все его свойства, включая стабильность. В среде ДУХ взаимодействие материальных частиц происходят в результате резонанса связанных с ними «облаков» среды (совпадения формы и частоты). Протон стабилен, так как защищён от материальных частиц и квантов оболочкой из пионов, имеющих иное волновое поле.
Масса протона 1836,153 me, а нейтрона 1838,683 me . Компенсацию заряда протона, по аналогии с атомом водорода, обеспечит электрон на волновой орбите в его экваториальной плоскости («одна ось вращения»), а его «двухосное вращение» оказывается «своим» в пионовом облаке. Добавим 2 бозона в противоположно расположенных пионах нейтрона; они компенсируют орбитальный момент, а масса нейтрона составит 1838,486 me. Такая структура объясняет массу нейтрона (отличие 0,01%), отсутствие заряда и, главное, - «ядерные» силы. «Лишний» бозон слабо связан в структуре и обеспечивает «обменную» связь, занимая с ядерной частотой «вакансию» в соседнем пионе протона, он вытесняет другой бозон, возвращающийся в нейтрон. «Лишние» бозоны в нейтроне - это его «две руки», скрепляющие ядра.
Нейтрон в ядрах элементов обеспечивает стабильность ядер, и сам «спасается» в ядре от распада (Т1/2 =11,7 мин.), причина которого - его «слабые места»: орбита электрона и наличие в «пионовой шубе» у двух из шести пионов по «лишнему» бозону.
Учёные ХХ века придумали десятки теорий и сотни «элементарных» частиц, но не смогли объяснить структуры атомов, а Природе потребовалось всего две подобных частицы, чтобы создать два нуклона, а из них 92 элемента и построить весь материальный МИР!!!
Альфа-структура атомных ядер
Изотопы всех элементов, наиболее распространенных в Природе, имеют чётное число нейтронов (исключение 4Be5 и 7N7). Всего из 291 стабильных изотопов 75 % имеет чётное число нейтронов и только 3 % чётно-нечётных ядер. Это свидетельствует о предпочтении связи протона с двумя нейтронами, отсутствии протон-протонных связей и «зарядовой независимости ядерных сил». Каркас ядер формируют связи нейтрон-протон, где каждый нейтрон может обменом двух бозонов удерживать 2 протона (пример, 2Не1). В тяжёлых ядрах относительное число нейтронов возрастает, усиливая каркас ядра.
Изложенные аргументы и принцип системности организации материи в не материальной среде позволяют предложить модель «блочного строительства» структуры ядер элементов, в которой «блоком» является ядро атома гелия - альфа-частица . Гелий - основной элемент космологического нуклеосинтеза, и по распространённости во Вселенной он второй элемент после водорода . Альфа-частицы являются оптимальной структурой прочно связанных двух пар нуклонов. Это очень компактная, крепко связанная шарообразная структура, которую геометрически можно представить как сферу с вписанным в неё кубом с узлами в противоположных диагоналях из 2 протонов и 2 нейтронов. Каждый нейтрон имеет две «ядерно-обменные» связи с двумя протонами. Электромагнитную связь сближения нейтрона с протонами обеспечивает орбитальный электрон в его структуре (подтверждение: магнитные моменты: μ (p) = 2,793 μN, μ (n) = -1,913 μN , где μN - ядерный магнетон Бора ).
Предполагаемое «кулоновское» отталкивание протонов не противоречит их сближению. Объяснение этому, также как в структурах мюонов из массонов, заложено в понимании «заряда» как неотъемлемого свойства массы частицы - движения среды ДУХ, связанного с волновым движением массы, выражающимся как сила в этой среде (единицей заряда может служить кулон2 - сила, умноженная на поверхность) . Два типа зарядов +/- - это левое и правое направление вращения. При сближении двух протонов в экваториальной плоскости движение «схваченной» среды будет противоположно, а при сближении «с полюсов» оно происходит в одном направлении, способствуя сближению. Сближение частиц ограничено взаимодействием их «полевых» оболочек, соответствующих «комптоновской» длине волны: λК(р) = 1,3214·10-15 м, а λК(n) = 1,3196·10-15 м. При сближении протона и нейтрона на такое расстояние действуют бозоно-обменные («ядерные») силы между ними.
Структуры ядер из альфа-частиц формируются с минимальным объёмом и формой, близкой к сферической. Структура альфа-частиц позволяет им объединяться путём разрыва одной бозоно-обменной связи n-p и формированием двух связей n-p и p-n с соседней альфа-частицей. При любом количестве протонов в ядре формируется единое сферическое поле, напряжённость которого такая же, как, если бы заряд был сосредоточен в центре (правило Остроградского - Гаусса). Образование единого поля ядра подтверждается орбитально-волновой структурой атомов, где все s, p, d, f орбиты образуют сферические оболочки .
Построение ядер элементов из альфа-частиц происходит системно, последовательно в каждом периоде на основе ядра предшествующего элемента. В ядрах с чётным числом протонов связи уравновешены, появление в структуре следующего атома дополнительного протона не возможно. В ядрах атомов после кислорода прибавление протона происходит по схеме (n-p-n). Чёткая последовательность формирования структур в соответствии с периодами и рядами в таблице Д.И. Менделеева - подтверждение правомерности предлагаемой модели ядер и служит подтверждением мысли В.И. Вернадского о «череде атомов»: «Процесс закономерной бренности атомов неизбежно и непреоборимо происходит… Беря историю любого атома в космическое время, мы видим, что он через определённые промежутки времени, сразу, одинаковыми скачками, в направлении полярного вектора времени переходит в другой атом, другой химический элемент» . Схемы ядер первых периодов атомов представлены в табл. 1.
Таблица 1
Предполагаемая структура ядер (плоская проекция) основных изотопов стабильных атомов из альфа-частиц (α), протонов (р) и нейтронов (n): pAn
|
nnααααααnn |
|||||||
|
nnααααααnn |
nnαααnnαααnn nnααnαααnααnn nαααnnαααn |
nnααααααnn nααnnαααnnααn nαααnnαααn |
Следующие 5 и 6 периоды элементов могут быть смоделированы аналогично с учётом того, что увеличение числа протонов потребует увеличения числа нейтронов как во внутреннем каркасе ядер, так в поверхностном слое, по схеме n-n.
Представленная наглядная плоская проекция структуры ядер может быть дополнена орбитальной схемой, соответствующей периодам в таблице Менделеева
(табл. 2).
Таблица 2
Ядерные оболочки элементов и периоды в таблице Д.И. Менделеева
|
Ядерная оболочка- период |
Начальный и конечный элемент в ряду |
Число элементов |
Отношение n/p |
|
|
Начальный |
Конечный |
|||
|
55Cs78 -82Pb126 (83Bi126… 86Rn136) |
||||
|
(87Fr136 - 92U146 …). |
||||
Оболочки строятся подобно структуре атома, где сферические оболочки из электронных орбит в каждом периоде формируются на большем радиусе, чем в предыдущем периоде .
Элементы после 82Pb126 (83Bi126 T1/2 ≈1018 лет) не стабильны (в табл. 2 даны в скобках). 41 альфа-частица в структуре свинца формируют электрический заряд, который для сохранения стабильности ядер требует силы дополнительных 40-44 нейтронов. Соотношение количества нейтронов и протонов n/p> (1,5÷1,6) - предел стабильности для тяжёлых ядер. Периоды полураспада ядер после 103 «элемента» - секунды. Эти «элементы» не могут сохранить структуру ядра и сформировать электронную оболочку атома. Вряд ли стоит тратить средства и время учёных на их искусственное производство. «Острова стабильности» быть не может!
Модель альфа-структуры ядер объясняет силы взаимосвязи, стабильность, и все свойства элементов (завершённость структуры инертных газов, распространённость в природе и особая стабильность элементов с симметричной структурой: О, С, Si, Mg, Ca, подобие Cu, Ag, Au…).
Причины «не спонтанного» распада
Структуры радиоактивных изотопов отличаются не симметричностью, наличием не уравновешенной пары n-p. Период полураспада изотопов тем меньше, чем больше их структура отличается от оптимальной. Радиоактивность изотопов с большим числом протонов объяснятся тем, что «обменные» силы нейтронов не способны удерживать их суммарный заряд, а распад изотопов с избытком нейтронов объясняется их излишеством для оптимальной структуры. Альфа-структура ядер позволяет объяснить причины всех видов радиоактивного распада .
Альфа-распад. В ядерной физике «согласно современным представлениям, альфа-частицы образуются в момент радиоактивного распада при встрече движущихся внутри ядра двух протонов и двух нейтронов… вылет альфа-частицы из ядра возможен благодаря туннельному эффекту через потенциальный барьер высотой не меньше 8,8 МэВ» . Всё происходит случайно: движение, встреча, формирование, набор энергии и вылет через некий барьер. В ядрах с альфа-структурой для вылета нет барьеров. Когда сила суммарного заряда всех протонов превышает бозоно-обменные силы сдерживания всех нейтронов, ядро сбрасывает альфа-частицу, наименее связанную в структуре, и «омолаживается» на 2 заряда. Появление возможности альфа-распада зависит от структуры ядер. Он проявляется при 31 альфа-частицt в ядре 62Sm84 (n/p =1,31), и становится необходимым от 84Ро (n/p = 1,48).
β+-распад. В ядерной физике «процесс β+- распада протекает так, как если бы один из протонов ядра превратился в нейтрон, испустив при этом позитрон и нейтрино: 11p→ 01n + +10e + 00νe… Так как масса протона меньше, чем у нейтрона, то такие реакции для свободного протона наблюдаться не могут. Однако, для протона, связанного в ядре, благодаря ядерному взаимодействию частиц, эти реакции оказываются энергетически возможными» . Объяснения процесса реакции, появления позитрона в ядре и увеличение массы на 2,5 me для превращения протона в нейтрон физика заменила постулатом: «процесс возможен». Такая возможность объясняется альфа-структурой. Рассмотрим классическую схему распада: 15Р15 → 14Si16 + +10e + 00νe. В соответствие с табл.1 структура стабильного изотопа 15Р16 (7α-npn). Cтруктура изотопа
15Р15 - (7α-np), но связь (n-p) в структуре - слабая, поэтому период полураспада 2,5 мин. Схема распада может быть представлена в несколько этапов. Слабо связанный протон выталкивается зарядом ядра, но «хватается» за нейтрон альфа-частицы и разрушает её с освобождением 4-х бозонов-связи. «Двухосные» бозоны не могут существовать в среде ДУХ и преобразуются в «трёхосные» массоны с разными моментами (+ и - ; электрон и позитрон) с испусканием нейтрино и антинейтрино по схемам
β-: (е--- + е+++ → е- -++ + ν0-) и β+: (е--- + е+++ → е+ --+ + ν0+). Позитрон выталкивается из ядра, а электрон на орбите вокруг бывшего протона компенсирует его заряд, превращая в нейтрон. Предполагаемая схема реакции: (7α-np) → (6α- n-p-n-р-n-p + 2е--- + 2e+++) → {(6 α) + (npnp) + n + (p-e-)} + e+ + ν0- + ν0+ → (7 α -nn) + e+ + ν0- + ν0+ . Схема объясняет причину и процесс распада, изменение массы частиц и предполагает испускание 2-х импульсов: нейтрино и антинейтрино.
β- -распад. «Поскольку электрон не вылетает из ядра и не вырывается из оболочки атома, было сделано предположение что β- электрон рождается в результате процессов, происходящих внутри ядра…» . Есть объяснение! Такой процесс характерен для ядер, имеющих в своей структуре количество нейтронов, большее, чем у стабильных изотопов этого элемента. Структура ядра следующего изотопа после ядра со сформированной чётно-чётной структурой прирастает «блоком» n-p-n, а следующий по массе за ним изотоп содержит ещё один «очень не лишний» нейтрон. Нейтрон может быстро «сбросить» орбитальный электрон, став протоном, и сформировать альфа-структуру: npn + (n→p) = npnp = α. Электрон и антинейтрино уносят избыток массы и энергии, а заряд ядра возрастает на единицу.
ε-захват. При недостатке нейтронов для стабильной структуры излишний заряд протонов притягивает и захватывает электрон с одной из внутренних оболочек атома, испуская нейтрино. Протон в ядре превращается в нейтрон.
Заключение
Представленная модель альфа-структуры ядер элементов позволяет объяснить закономерности образования ядер, их стабильность, причины, стадии и энергетический баланс всех видов радиоактивного распада. Структуры протонов, нейтронов, ядер и атомов элементов, подтверждённые соответствием универсальным постоянным, которые являются физическими характеристиками среды ДУХ, объясняют все свойства и все взаимодействия. Современная ядерная и атомная физика на это не способны. Необходим пересмотр основных концепций: от постулатов - к пониманию.
Библиографическая ссылка
Поляков В.И. СТРУКТУРА ЯДЕР АТОМОВ И ПРИЧИНЫ РАДИОАКТИВНОСТИ // Успехи современного естествознания. – 2014. – № 5-2. – С. 125-130;URL: http://natural-sciences.ru/ru/article/view?id=33938 (дата обращения: 27.02.2019). Предлагаем вашему вниманию журналы, издающиеся в издательстве «Академия Естествознания»
Введение
Глава 1. Структура и химия клеточного ядра. Открытие ядра. Роберт Броун
1.1. Интерфазное ядро
1.2. Работы флемминга
1.3. Ядрышки
1.4. Ядерная мембрана
1.5. Кариоплазма
1.6. Хроматин
Глава 2. Клеточное ядро - центр управления жизнедеятельностью клетки
2.1. Ядро - незаменимый компонент клетки
2.2. Функциональная структура ядра
2.3. Роль ядерных структур в жизнедеятельности клетки
2.4. Ведущее значение днк
Список литературы
ВВЕДЕНИЕ
Клеточное ядро - центр управления жизнедеятельностью клетки. Из общей схемы белкового синтеза можно видеть, что начальным пунктом, с которого начинается поток информации для биосинтеза белков в клетке, является ДНК. Следовательно, именно ДНК содержит ту первичную запись информации, которая должна сохраняться и воспроизводиться от клетки к клетке, из поколения в поколение. Кратко касаясь вопроса о месте хранения генетической информации, т. е. о локализации ДНК в клетке, можно сказать следующее. Уже давно известно, что, в отличие от всех прочих компонентов синтезирующего белок аппарата, универсально распределенных по всем частям живой клетки, ДНК имеет особую, весьма ограниченную локализацию: местом ее нахождения в клетках высших (эукариотических) организмов является клеточное ядро.
У низших (прокариотических) организмов, не имеющих оформленного клеточного ядра, - бактерий и сине-зеленых водорослей, - ДНК также отделена от остальной части протоплазмы одним или несколькими компактными нуклеоидными образованиями. В полном соответствии с этим ядро эукариотов или нуклеоид прокариотов издавна рассматриваются как вместилище генов, как уникальный клеточный органоид, контролирующий реализацию наследственных признаков организмов и их передачу в поколениях. Генетические данные о «единоначалии» ядра в клетке всегда непосредственно объединялись с биохимическими данными об уникальной локализации ДНК в ядре.
1. СТРУКТУРА И ХИМИЯ КЛЕТОЧНОГО ЯДРА. ОТКРЫТИЕ ЯДРА. РОБЕРТ БРОУН
Термин «ядро» впервые был применен Броуном в 1833 г. для обозначения шаровидных постоянных структур в клетках растений. В 1831-1833 гг., шотландский путешественник и физик (открывший «броуновское движение») Роберт Броун (1773-1858) обнаружил ядро в растительных клетках. Он дал ему название «Nucleus», или «Areola». Первый термин стал общепринятым и сохранился по настоящее время, второй же распространения не получил и забыт. Весьма важно, что Броун настаивал на постоянном наличии ядра во всех живых клетках.
Роль и значение клеточного ядра не были в то время известны. Полагали, что оно представляет собой «конденсированную в комочек слизь, а возможно, и запасное питательное вещество». Позднее такую же структуру описали во всех клетках высших организмов. Говоря о клеточном ядре, мы имеем в виду собственно ядра эукариотических клеток. Их ядра построены сложным образом и довольно резко отличаются от «ядерных» образований, нуклеоидов прокариотических организмов. У последних в состав нуклеоидов (ядроподобных структур) входит одиночная, кольцевая молекула ДНК, практически лишенная белков. Иногда такую молекулу ДНК бактериальных клеток называют бактериальной хромосомой, или генофором (носителем генов).
Бактериальная хромосома не отделена мембранами от основной цитоплазмы, однако собрана в компактную, ядерную зону, нуклеоид, который можно видеть в световом микроскопе после специальных окрасок или же в электронном микроскопе. Анализируя структуру и химию клеточного ядра, мы будем опираться на данные, касающиеся ядер эукариотических клеток, постоянно сравнивая их с ядрами прокариотов. Клеточное ядро, обычно одно на клетку (есть примеры многоядерных клеток), состоит из ядерной оболочки, отделяющей его от цитоплазмы, хроматина, ядрышка и кариоплазмы или ядерного сока. Эти четыре основных компонента встречаются практически во всех неделящихся клетках эукариотических одно- или многоклеточных организмов.
1.2. РАБОТЫ ФЛЕММИНГА
До некоторых пор роль ядра в клеточном делении оставалась неопределенной. Это, вероятно, было связано с трудностью наблюдения за ним. В живой клетке ядро, как правило, можно видеть только при значительном увеличении обычного светового микроскопа. Ядро, находящееся в процессе деления, наблюдать еще труднее. Анилиновые красители окрашивают ядро, цитоплазму и клеточную оболочку по-разному и, следовательно, облегчают узнавание этих структур.
Анилиновые красители синтезируются искусственно, и методика их получения не была известна до середины XIX в. Естественные красители, которые биологи использовали раньше, не всегда окрашивали ядра достаточно хорошо, чтобы их можно было отличить от остальных частей клетки. И вновь дальнейший прогресс зависел от развития подходящих для проведения исследований методов. В то время не было недостатка в хороших микроскопах, но не было известно, как обрабатывать клетки, чтобы увидеть как можно больше клеточных структур. Следует отметить, что никто не знал, будут ли анилиновые красители для этой цели лучше, чем естественные.
Когда в 1860-х гг. химики получили анилиновые красители, кто-то просто наугад попытался использовать их для окрашивания тонких срезов растительных и животных тканей. В 1879 г. немецкий биолог Вальтер Флемминг использовал различные анилиновые красители и ахроматические линзы. Обработав клетки красителями и изучая их под микроскопом с ахроматическими линзами, он проследил за поведением ядра в процессе клеточного деления. В его книге «Клеточное вещество, ядро и клеточное деление» описаны результаты наблюдений над клеточным делением, причем описания очень близки к современным.
Поскольку хромосомы похожи на нити, Флемминг решил назвать этот процесс митозом (греческое слово, что в переводе значит «нить»). Строго говоря, митоз относится только к процессу ядерного удвоения. Образования клеточной пластинки в растительных клетках и клеточной бороздки в животных клетках являются делениями цитоплазмы.
Было бы неправильным считать, что Флемминг - единственный первооткрыватель явления митоза. Понимание всей последовательности процесса митоза зависело от многих ученых, работавших над этой проблемой все предыдущие годы. Одна из основных трудностей исследования событий, происходящих в клетке, состояла в том, что клетки погибали в процессе окрашивания. Это означает, что клетка изучается только после того, как жизнедеятельность в ней прекращена. По этой «остановленной в движении» картине Флемминг и другие исследователи воссоздали то, что происходит в живых клетках. Это примерно то же, что воссоздать работу фабрики по серии моментальных снимков, взятых в различные интервалы времени. По существу, это и было сделано Флеммингом. Другие ученые, основываясь на работе Флеминга, в конце концов выявили связь хромосом с наследственностью и эволюцией.
Именно так развивается наука: успех зависит не от случайных открытий ученых-«гигантов», а от кропотливой работы большого отряда ученых. В световом, а также в фазово-контрастном микроскопах ядро обычно представляется оптически гомогенным: видны лишь оболочка и одно или несколько ядрышек внутри. Иногда обнаруживаются также гранулы и небольшие глыбки. Реже в неделящихся живых клетках удается наблюдать хромосомы. Тонкая хроматиновая сеть отчетливо выявляется лишь после фиксации и окрашивания клетки основными красителями.
Исследования ядра на фиксированных и окрашенных препаратах показали, что его микроскопическое изображение почти не зависит от метода изготовления препаратов. Лучше всего тонкая структура ядра сохраняется при фиксации четырехокисью осмия. Другие общепринятые фиксаторы позволяют различать на препарате ядерную оболочку, ядрышко, хроматиновые структуры в виде глыбок и нитей и неокрашенную массу между ними - нуклеоплазму.
Хроматиновые структуры расположены в более жидкой ахроматической среде, они могут быть плотными или рыхлыми, пузыревидными. У некоторых объектов хроматин после фиксации не образует явно выраженной ядерной сети, а концентрируется в ядре в виде крупных глыбок, названных хромоцентрами, или прохромосомами. В ядрах подобного типа весь хроматин сосредоточен в хромоцентрах.
1.3. ЯДРЫШКИ
Согласно электронно-микроскопическим исследованиям, ядрышки лишены какой-либо мембраны. Вещество их в основном состоит из субмикроскопических нитей и нуклеоплазмы. Ядрышки можно наблюдать, применяя специальные методы окрашивания, а также в ядрах некоторых живых клеток при использовании фазово-контрастного микроскопа или темнопольного конденсора.
На электронных микрофотографиях в ядрышках нередко видны две зоны: центральная - гомогенная и периферическая - построенная из гранулированных нитей. Эти гранулы напоминают рибосомы, но отличаются от них меньшей плотностью и величиной. Ядрышки богаты белками (80-85 %) и РНК (около 15 %) и служат активными центрами синтеза рибосомальной РНК. В соответствии с этим главной составной частью ядрышка является ядрышковая ДНК, которая принадлежит организатору ядрышек одной из хромосом.
содержание РНК заметно колеблется, в зависимости от интенсивности обмена веществ в ядре и цитоплазме. Ядрышки не присутствуют в ядре постоянно: они возникают в средней телофазе митоза и исчезают в конце профазы. Полагают, что по мере затухания синтеза РНК в средней профазе происходят разрыхление ядрышка и выход в цитоплазму образовавшихся в нуклеоплазме субчастиц рибосом. При исчезновении ядрышка во время митоза его белки, ДНК и РНК, становятся основой матрикса хромосом, а в дальнейшем из материала старого ядрышка формируется новое.
Установлена связь ядрышек с хромосомами, имеющими спутников, поэтому число ядрышек соответствует числу спутничных хромосом. Нуклеолонемы сохраняются на протяжении всего цикла клеточного деления и в телофазе переходят от хромосом к новому ядрышку.
1.4. ЯДЕРНАЯ МЕМБРАНА
Неделящееся клеточное ядро заключено в плотную и упругую оболочку, которая растворяется и вновь восстанавливается в процессе деления клетки. Это образование отчетливо видно лишь на некоторых объектах, например у гигантских ядер слизевых клеток алоэ толщина мембраны достигает 1 мкм. В световом микроскопе структуру ядерной оболочки удается наблюдать лишь у плазмолизированных клеток, фиксированных и окрашенных.
Детальное изучение ядерной мембраны стало возможным с появлением электронной микроскопии. Исследования показали, что наличие ядерной оболочки характерно для всех эукариотических клеток. Она состоит из двух элементарных мембран толщиной 6-8 нм каждая - внешней и внутренней, между которыми находится перинуклеарное пространство шириной от 20 до 60 нм. Оно заполнено энхилемой - сывороткообразной жидкостью с низкой электронной плотностью.
Итак, ядерная мембрана представляет собой полый мешок, отделяющий содержимое ядра от цитоплазмы, и состоит из двух слоев: внешний слой ограничивает перинуклеарное пространство снаружи, т. е. со стороны цитоплазмы, внутренний - изнутри, т. е. со стороны ядра. Из всех внутриклеточных мембранных компонентов подобным строением мембран обладают ядро, митохондрии и пластиды.
Морфологическое строение каждого слоя такое же, как и внутренних мембран цитоплазмы. Отличительная особенность ядерной оболочки - наличие в ней пор - округлых перфораций, образующихся в местах слияния внешней и внутренней ядерных мембран. Размеры пор довольно стабильны (30-100 нм в диаметре), в то же время их число изменчиво и зависит от функциональной активности клетки: чем активнее идут в ней синтетические процессы, тем больше пор приходится на единицу поверхности клеточного ядра.
Обнаружено, что количество пор увеличивается в период реконструкции и роста ядра, а также при репликации ДНК. Одно из крупнейших открытий, сделанных с помощью электронной микроскопии, - обнаружение тесной взаимосвязи между ядерной оболочкой и эндоплазматической сетью. Поскольку ядерная оболочка и тяжи эндоплазматической сети во многих местах сообщаются между собой, перинуклеарное пространство должно содержать ту же сывороткообразную жидкость, что и полости между мембранами эндоплазматической сети.
При оценке функциональной роли ядерной оболочки большое значение приобретает вопрос о ее проницаемости, обусловливающей обменные процессы между ядром и цитоплазмой в связи с передачей наследственной информации. Для правильного понимания ядерно-цитоплазматических взаимодействий важно знать, насколько ядерная оболочка проницаема для белков и других метаболитов. Опыты показывают, что ядерная оболочка легко проницаема для относительно крупных молекул. Так, рибонуклеаза - фермент, гидролизующий рибонуклеиновую кислоту без выделения свободной фосфорной кислоты, - имеет молекулярную массу около 13000 и очень быстро проникает в ядро.
Даже в корешках, фиксированных видоизмененным методом замораживания, можно наблюдать, как окрашивание ядрышек подавляется во всех клетках уже через 1 ч после обработки рибонуклеазой.
1.5. КАРИОПЛАЗМА
Кариоплазма (ядерный сок, нуклеоплазма) - основная внутренняя среда ядра, она занимает все пространство между ядрышком, хроматином, мембранами, всевозможными включениями и другими структурами. Кариоплазма под электронным микроскопом имеет вид гомогенной или мелкозернистой массы с низкой электронной плотностью. В ней во взвешенном состоянии находятся рибосомы, микротельца, глобулины и различные продукты метаболизма.
Вязкость ядерного сока примерно такая же, как вязкость основного вещества цитоплазмы. Кислотность ядерного сока, определенная путем микроинъекции индикаторов в ядро, оказалась несколько выше, чем у цитоплазмы.
Кроме того, в ядерном соке содержатся ферменты, участвующие в синтезе нуклеиновых кислот в ядре и рибосомы. Ядерный сок не окрашивается основными красителями, поэтому его называют ахроматиновым веществом, или кариолимфой, в отличие от участков, способных окрашиваться, - хроматина.
1.6. ХРОМАТИН
Термин «хромосома» используется по отношению к молекуле нуклеиновой кислоты, которая представляет собой хранилище генетической информации вируса, прокариота или эукариотической клетки. Однако первоначально слово «хромосома» (т. е. «окрашенное тело») использовалось в другом смысле, - для обозначения густо окрашенных образований в эукариотических ядрах, которые можно было наблюдать в световой микроскоп после обработки клеток красителем.
Эукариотические хромосомы, в изначальном смысле этого слова, выглядят как резко очерченные структуры только непосредственно до и во время митоза - процесса деления ядра в соматических клетках. В покоящихся, неделящихся эукариотических клетках хромосомный материал, называемый хроматином, выглядит нечетко и как бы беспорядочно распределен по всему ядру. Однако, когда клетка готовится к делению, хроматин уплотняется и собирается в свойственное данному виду число хорошо различимых хромосом.
Хроматин был выделен из ядер и проанализирован. Он состоит из очень тонких волокон, которые содержат 60 % белка, 35 % ДНК и, вероятно, 5 % РНК. Хроматиновые волокна в хромосоме свернуты и образуют множество узелков и петель. ДНК в хроматине очень прочно связана с белками, называемыми гистонами, функция которых состоит в упаковке и упорядочении ДНК в структурные единицы - нуклеосомы. В хроматине содержится также ряд негистоновых белков. В отличие от эукариотических, бактериальные хромосомы не содержат гистонов; в их состав входит лишь небольшое количество белков, способствующих образованию петель и конденсации (уплотнению) ДНК.
Глава 2. КЛЕТОЧНОЕ ЯДРО - ЦЕНТР УПРАВЛЕНИЯ ЖИЗНЕДЕЯТЕЛЬНОСТЬЮ КЛЕТКИ
2.1. ЯДРО - НЕЗАМЕНИМЫЙ КОМПОНЕНТ КЛЕТКИ
Еще в конце прошлого века было доказано, что лишенные ядра фрагменты, отрезанные от амебы или инфузории, через более или менее короткое время погибают. Более детальные опыты показали, что энуклеированные амебы живут, но вскоре после операции перестают питаться, двигаться и через несколько дней (до одной недели) погибают. Если пересадить ядро в ранее энуклеированную клетку, то процессы нормальной жизнедеятельности восстанавливаются и через некоторое время амеба начинает делиться.
Яйцеклетки морского ежа, лишенные ядра, при стимуляции к партеногенетическому развитию делятся, но тоже в конце концов погибают. Особенно интересные опыты были проведены на крупной одноклеточной водоросли ацетабулярии. После удаления ядра водоросль не только живет, но и в течение определенного периода может восстанавливать безъядерные участки. Следовательно, при отсутствии ядра прежде всего нарушается способность к размножению, и, хотя жизнеспособность на какое-то время сохраняется, в конце концов такая клетка неизбежно погибает.
содержание ядерного и лишенного ядра фрагмента в среде с радиоактивным предшественником РНК - 3Н-уридином показало, что синтез РНК в безъядерном фрагменте отсутствует. Белковый же синтез продолжается некоторое время за счет информационных РНК и рибосом, сформированных ранее, до удаления ядра. Пожалуй, наиболее яркую иллюстрацию роли ядра дают безъядерные эритроциты млекопитающих. Это эксперимент, поставленный самой природой.
Созревая, эритроциты накапливают гемоглобин, затем выбрасывают ядро и в таком состоянии живут и функционируют в течение 120 дней. Они не способны размножаться и в конце концов погибают. Однако клетки, только что выбросившие ядро, так называемые ретикулоциты, еще продолжают синтез белка, но уже не синтезируют РНК. Следовательно, удаление ядра влечет за собой прекращение поступления в цитоплазму новых РНК, которые синтезируются на молекулах ДНК, локализованных в хромосомах ядра. Однако это не мешает уже существующей в цитоплазме информационной РНК продолжать синтезировать белок, что и наблюдается в ретикулоцитах. Затем, когда РНК распадается, синтез белка прекращается, но эритроцит еще продолжает жить долгое время, выполняя свою функцию, которая не связана с интенсивным расходованием белка.
Лишенные ядра яйцеклетки морского ежа продолжают жить и могут делиться благодаря тому, что во время овогенеза запасли значительное количество РНК, которая и продолжает функционировать. Информационная РНК у бактерий функционирует минуты, но в ряде специализированных клеток млекопитающих она сохраняется сутки и больше.
Несколько особняком стоят данные, полученные на ацетобулярии. Оказалось, что морфогенез удаленной части определяется ядром, но жизнь кусочка обеспечивается ДНК, которую содержат хлоропласты. На этой ДНК синтезируется информационная РНК, которая, в свою очередь, обеспечивает синтез белка.
2.2. ФУНКЦИОНАЛЬНАЯ СТРУКТУРА ЯДРА
В изучении структурно-биохимической организации ядерного аппарата различных клеток большую роль играют сравнительно-цитологические исследования, в которых применяются как традиционный эволюционно-исторический подход, так и широкие сравнительно-цитологические сопоставления организации ядерного аппарата различных разновидностей клеток. Эволюционно-историческое направление в этих исследованиях имеет особое значение, поскольку ядерный аппарат представляет собой наиболее консервативную клеточную структуру - структуру, ответственную за хранение и передачу генетической информации.
Широкое сравнительно-цитологическое изучение ядерного аппарата у тех клеток, которые как бы резко уклоняются от обычного (типичного) уровня организации (ооциты, сперматозоиды, ядерные эритроциты, инфузории и т. д.), и использование данных, полученных с помощью молекулярно-биологических и цитологических методов в специальных науках, занимающихся клеточным уровнем организации (частная цитология, протозоология и т. д.), позволили выявить массу интересных особенностей организации ядерного аппарата, имеющих общецитологическое значение.
В составе ядерного аппарата эукариотных клеток можно выделить ряд субсистем, центральное место среди которых занимает совокупность интерфазных хромосом, или ДНК ядра. В них сосредоточена вся ДНК ядра, находящаяся в весьма сложных взаимоотношениях с белками хроматина, которые, в свою очередь, подразделяются на структурные, функциональные и регуляторные белки.
Второй и весьма важной субсистемой ядерного аппарата является ядерный матрикс, представляющий собой систему фибриллярных белков, выполняющих как структурную (скелетную) функцию в топографической организации всех ядерных компонентов, так и регуляторную функцию в организации процессов репликации, транскрипции, в созревании (процессинге) и перемещении продуктов транскрипции внутри ядра и за его пределы. По-видимому, белковый матрикс имеет двоякую природу: какие-то одни его компоненты обеспечивают в основном скелетную функцию, другие - регуляторную и транспортную.
Вместе с определенными участками ДНК хроматина белки ядерного матрикса (функционального и структурного) образуют основу ядрышка. Белки структурного матрикса принимают участие и в формировании поверхностного аппарата ядра. Поверхностный аппарат ядра занимает и в структурном, и в функциональном отношениях промежуточное положение между метаболическим аппаратом цитоплазмы и ядром. Мембраны и цистерны ядерной оболочки являются по сути дела специализированной частью общей мембранной системы цитоплазмы.
Специфическими структурами поверхностного аппарата ядра, играющими важную роль в реализации его основной функции - обеспечении взаимодействия ядра и цитоплазмы выступают поровые комплексы и субмембранная плотная пластинка, которые образуются с помощью белков ядерного матрикса. Наконец, последней субсистемой ядерного аппарата является кариоплазма. Это аналогичная гиалоплазме внешне бесструктурная фаза ядерного аппарата, которая создает специфическое для ядерных структур микроокружение, что обеспечивает возможность их нормального функционирования.
Кариоплазма находится в постоянном взаимодействии с гиалоплазмой через систему поровых комплексов и мембран ядерной оболочки.
2.3. РОЛЬ ЯДЕРНЫХ СТРУКТУР В ЖИЗНЕДЕЯТЕЛЬНОСТИ КЛЕТКИ
Основные процессы, связанные с синтезом белка, в принципе одинаковы у всех форм живого, указывают на особое значение клеточного ядра. Ядро осуществляет две группы общих функций: одну, направленную на собственно хранение генетической информации, другую - на ее реализацию, на обеспечение синтеза белка. Иными словами, первую группу составляют процессы поддержания наследственной информации в виде неизменной структуры ДНК. Эти процессы обусловлены наличием так называемых репарационных ферментов, ликвидирующих спонтанные повреждения молекул ДНК (разрыв одной из цепей ДНК, часть радиационных повреждений), что сохраняет строение молекул ДНК практически неизменными в ряду поколений клеток или организмов.
Далее в ядре происходит воспроизведение, или редупликация, молекул ДНК, что дает возможность двум клеткам получить совершенно одинаковые и в качественном, и в количественном смысле объемы генетической информации. В ядрах происходят процессы изменения и рекомбинации генетического материала, что наблюдается во время мейоза (кроссинговер). Наконец, ядра непосредственно участвуют в процессах распределения молекул ДНК при делении клеток.
Другой группой клеточных процессов, обеспечивающихся активностью ядра, является создание собственно аппарата белкового синтеза. Это не только синтез, транскрипция на молекулах ДНК разных информационных РНК, но транскрипция всех видов трансферных РНК и рибосомных РНК. В ядре эукариотов происходит также образование субъединиц рибосом путем комплексирования синтезированных в ядрышке рибосомных РНК с рибосомными белками, которые синтезируются в цитоплазме и переносятся в ядро. Таким образом, ядро представляет собой не только вместилище генетического материала, но и место, где этот материал функционирует и воспроизводится. Поэтому выпадение или нарушение любой из перечисленных выше функций гибельно для клетки в целом.
Так, нарушение репарационных процессов будет приводить к изменению первичной структуры ДНК и автоматически - к изменению структуры белков, что непременно скажется на их специфической активности, которая может просто исчезнуть или измениться так, что не сможет обеспечивать клеточные функции, в результате чего клетка погибает. Нарушения редупликации ДНК приведут к остановке размножения клеток или к появлению клеток с неполноценным набором генетической информации, что тоже гибельно для них. К такому же результату приведет нарушение процессов распределения генетического материала (молекул ДНК) при делении клеток.
Выпадение в результате поражения ядра или в случаях нарушений каких-либо регуляторных процессов синтеза любой формы РНК автоматически приведет к остановке синтеза белка в клетке или к грубым его нарушениям. Все это указывает на ведущее значение ядерных структур в процессах, связанных с синтезом нуклеиновых кислот и белков, главных функционеров в жизнедеятельности клетки.
Ядро осуществляет сложную координацию и регуляцию процессов синтеза РНК. Как указывалось, все три типа РНК образуются на ДНК. Радиографическими методами показано, что синтез РНК начинается в ядре (хроматине и ядрышке), и уже синтезированная РНК перемещается в цитоплазму. Таким образом мы видим, что ядро программирует синтез белка, который осуществляется в цитоплазме. Однако само ядро также испытывает влияние цитоплазмы, т. к. синтезируемые в ней ферменты поступают в ядро и необходимы для его нормального функционирования. Например, в цитоплазме синтезируется ДНК-полимераза, без которой не может происходить авторепродукция молекул ДНК. Поэтому следует говорить о взаимном влиянии ядра и цитоплазмы, при котором главенствующая роль все же принадлежит ядру как хранителю наследственной информации, которая передается при делении от одной клетки к другой.
2.4. ВЕДУЩЕЕ ЗНАЧЕНИЕ ДНК
Основное биологическое значение ядерного аппарата определяется его главным компонентом - гигантскими молекулами ДНК, способными к репликации и транскрипции. Эти два свойства ДНК и лежат в основе двух важнейших функций ядерного аппарата любой клетки:
а) удвоения наследственной информации и передачи ее в ряду клеточных поколений;
б) регулируемой транскрипции участков молекул ДНК и транспорта синтезируемых РНК в цитоплазму клеток.
По характеру организации ядерного аппарата все клетки делятся на три группы: прокариотные, мезокариотные и эукариотные.
Клеткам прокариот свойственны отсутствие ядерной оболочки, укладка ДНК без участия гистонов, унирепликонный тип репликации ДНК, моноцистронный принцип организации транскрипции и ее регуляция преимущественно по принципу положительной и отрицательной обратной связи.
Клетки эукариот, напротив, отличаются наличием ядерной оболочки, точнее говоря, даже сложного поверхностного аппарата ядра и мультирепликонным типом репликации молекул ДНК, образующих набор хромосом. Упаковка этих молекул происходит с помощью комплекса белков. Характер упаковки подвергается циклическим изменениям, связанным с прохождением клетками закономерных фаз цикла репродукции. Процессы транскрипции ДНК и ее регуляции у эукариот значительно отличаются от таковых у прокариот.
Мезокариотные клетки по организации ядерного аппарата занимают как бы промежуточное положение между эукариотными и прокариотными клетками. У мезокариот, как и у эукариот, имеется хорошо развитый поверхностный аппарат ядра. Укладка в хромосомы молекул ДНК существенно отличается от организации ДНП в эукариотных клетках. Механизмы репликации и транскрипции ДНК у мезокариот выяснены слабо. Таким образом в клеточном ядре протекают важнейшие процессы, связанные с наследственным статусом организма, - peпликация (биосинтез ДНК) и транскрипция.
Кроме того, ядро является источником отдельных белков и ферментов, необходимых для жизнедеятельности дифференцированных тканей. Одновременно с потоком информации в клетку для обеспечения синтеза белков осуществляется обратная связь: цитоплазма - ядро, т. е. ядро функционирует в тесном взаимодействии с другими частями клетки, объединяя процессы ядерно-цитоплазматического транспорта и регуляторного взаимодействия с цитоплазмой клетки.
ЯДЕРНАЯ СТРУКТУРА АТОМА
Альфа-частицы . В 1896 году французским физиком Беккерелем было открыто явление радиоактивности. После этого начался быстрый прогресс в изучении строения атома. Этому, прежде всего способствовало то, что в руках физиков оказался очень эффективный инструмент исследования атомной структуры – α -частица. С помощью α -частиц, испускаемых естественными радиоактивными веществами, были сделаны важнейшие открытия: установлена ядерная структура атома, получены первые ядерные реакции, обнаружено явление искусственной радиоактивности и, наконец, найден нейтрон, сыгравший важную роль как при объяснении строения атомного ядра, так и при открытии процесса деления тяжелых ядер.
Альфа-частицы – это движущиеся с большой скоростью ядра гелия. Измерения скоростей α- частиц естественных излучателей по отклонению в электрическом и магнитном полях дали величину скорости (1,5-2).10 7 м/с, что соответствует кинетической энергии 4,5-8 МэВ (1 МэВ=1,6.10 -13 Дж). Такие частицы движутся в веществе прямолинейно, быстро теряют энергию на ионизацию атомов и после остановки превращаются в нейтральные атомы гелия.
Рассеяние альфа-частиц. Опыты Резерфорда. Изучая прохождение коллимированного пучка альфа-частиц через тонкую металлическую фольгу, английский физик Резерфорд обратил внимание на размытие изображения пучка частиц на регистраторе – фотопластинке. Резерфорд объяснил это размытие рассеянием альфа-частиц. Детальное изучение рассеяния альфа-частиц показало, что в редких случаях они рассеиваются на большие углы иногда превышающие 90 0 , что соответствует отбрасыванию быстродвижущихся частиц в обратном направлении. Такие случаи рассеяния в рамках модели Томпсона объяснить нельзя.
Тяжелая альфа-частица в одном акте столкновения может быть отброшена назад только при взаимодействии с частицей большей массы, превосходящей массу альфа-частицы. Электроны не могут быть такими частицами. Кроме того, рассеяние в обратном направлении предполагает сильное торможение альфа-частицы, т.е. энергия взаимодействия должна быть порядка кинетической энергии альфа-частицы. Энергия же электростатического взаимодействия альфа-частицы с атомом Томпсона, имеющим положительный заряд, распределенный в объеме или на поверхности атома радиусом 10 -8 см и равный в единицах элементарного заряда примерно половине атомной массы, много меньше этой величины. Результаты опыта можно объяснить, если расстояние от альфа частицы до центра положительного электрического заряда порядка 10 -12 см. Такое расстояние в 10000 раз меньше радиуса атома, а радиус положительного заряда должен быть еще меньше. Предположение о малом объеме рассеивающего центра согласуется с очень малым количеством случаев рассеяния на большие углы.
Для объяснения результатов своих наблюдений по рассеянию альфа-частиц Резерфорд предложил ядерную модель атома . Согласно этой модели в центре атома находится ядро, занимающее очень малый объем, содержащее почти всю массу атома и несущее положительный электрический заряд. Основной объем атома занят движущимися электронами, число которых равно числу элементарных положительных зарядов ядра, т.к. атом в целом нейтрален.
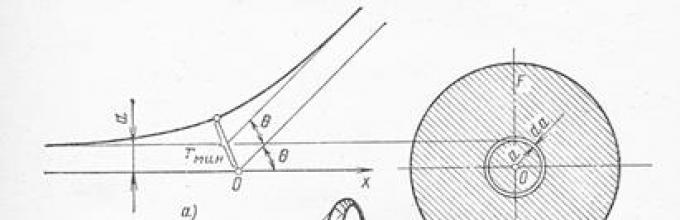
Теория рассеяния альфа-частиц. Чтобы обосновать предположение о ядерной структуре атома и доказать, что рассеяние альфа-частиц происходит в результате кулоновского взаимодействия с ядром, Резерфорд развил теорию рассеяния альфа-частиц точечными электрическими зарядами с большой массой и получил зависимость между углом рассеяния θ и числом частиц, рассеянных на угол θ . Если альфа-частица движется в направлении точечного заряда Ze , где Z – число элементарных зарядов, и при этом ее начальная траектория отстоит от оси, проходящей через рассеивающий центр, на расстоянии а (рис.1.1), то на основании закона Кулона методами классической механики можно вычислить угол θ , на который отклонится альфа-частица вследствие электростатического отталкивания одноименных электрических зарядов:
где M и v – масса и скорость альфа-частицы; 2e – ее заряд; ε 0 – электрическая постоянная, равная 8,85.10 -12 Ф/м.

Рис.1.1. Рассеяние альфа-частицы электрическим полем атомного ядра:
а) – схема рассеяния в плоскости траектории частицы; б) – кольцо, из которого происходит рассеяние под углом θ ; в) – схема рассеяния в конический телесный угол под угломθ к оси.
Доля частиц dn/n 0 , имеющих параметр удара а , от полного числа n 0 , падающих на мишень, равна доле элементарной площадки 2πada на полной площади F поперечного сечения пучка альфа-частиц (рис. 1.1, б). Если на площади F имеется не один, а N F рассеивающих центров, то соответствующая доля возрастет в N F раз и отнесенная к единице а, составит:
![]() , (1.2)
, (1.2)
где N 1 – число рассеивающих центров на единице площади мишени.
Учитывая, что dΩ=2π sinθ dθ, можно получить долю частиц, рассеянных в единицу конического телесного угла под углом θ к оси, как :
 (1.3)
(1.3)
Экспериментальная проверка полностью подтвердила последнюю зависимость при рассеянии веществом альфа-частиц. Строгое выполнение закона 1/sin 4
свидетельствует о том, что за рассеяние ответственны только электрические силы и что геометрические размеры электрических зарядов обоих тел по крайней мере меньше наименьшего расстояния в акте рассеяния r мин
. Расстояние r мин
тем меньше, чем больше угол рассеяния θ
. При θ
=π () оно самое малое и определяется условием ![]() , которое соответствует случаю обращения всей кинетической энергии альфа-частицы в потенциальную энергию отталкивания одноименных зарядов.
, которое соответствует случаю обращения всей кинетической энергии альфа-частицы в потенциальную энергию отталкивания одноименных зарядов.
По результатам обработки результатов экспериментов, исходя из различных в то время оценок величины заряда ядер Z , Резерфорд оценил радиус ядра величиной порядка 10 -12 см.
Атом Резерфорда-Бора. С открытием атомного ядра возникла необходимость объяснения устойчивости атома. С точки зрения классической электродинамики атом Резерфорда не может существовать длительно. Поскольку разноименные заряды притягиваются, электроны могут находится на определенном расстоянии от ядра только при условии движения вокруг ядра. Однако движение по замкнутой траектории является движением с ускорением, а движущийся с ускорением электрический заряд излучает энергию в окружающее пространство. За ничтожно малое время любой атом должен излучить энергию движения электронов и уменьшиться до размера ядра.
Первую стационарную модель атома предложил датский физик Нильс Бор в 1913 году. Бор связал устойчивость атомов с квантовой природой излучения. Гипотеза квантов энергии, выдвинутая немецким физиком Планком в 1900 году для объяснения спектра излучения абсолютно черного тела, утверждала, что микроскопические системы способны излучать энергию лишь определенными порциями – квантами с частотой v , пропорциональной энергии кванта E:
где h – универсальная постоянная Планка, равная 6,62.10 -24 Дж.с.
Бор предположил, что энергия атомного электрона в кулоновском поле ядра не меняется непрерывно, а принимает ряд устойчивых дискретных значений, которым соответствуют стационарные орбиты электронов. При движении по таким орбитам электрон не излучает энергию. Излучение атома происходит только при переходе электрона с орбиты с более высоким значением энергии на другую стационарную орбиту. Это излучение характеризуется единственным значением частоты, пропорциональной разности энергий орбит:
hv=Е нач - Е кон
Условие стационарности орбиты – это равенство механического момента количества движения электрона целому кратному числу h/2π :
mvr n = n ,
где mv – модуль импульса электрона;
r n – радиус n -ой стационарной орбиты;
n – любое целое число.
Введенное Бором условие квантования круговых орбит позволило рассчитать спектр атома водорода и вычислить спектроскопическую константу Ридберга для атома водорода. Система уровней одноэлектронного атома и радиусы стационарных орбит можно определить из последнего соотношения и закона Кулона:
![]() ; (1.4)
; (1.4)
Расчет по этим формулам при n=1 и Z=1 дает радиус наименьшей стационарной орбиты электрона в атоме водорода или первый боровский радиус:
![]() . (1.6)
. (1.6)
Движение электрона по орбите можно представить как замкнутый электрический ток и вычислить создаваемый им магнитный момент. Для первой орбиты водорода он называется магнетоном Бора и равен:
![]() (1.7)
(1.7)
Магнитный момент обратно пропорционален массе частицы, но для частиц данного сорта, например электронов, имеет смысл единицы. Характерно, что как раз этой единице равен собственный момент электрона, связанный с его спином.
Ядерная модель атома с электронами на устойчивых орбитах называется планетарной моделью Резерфорда-Бора. Она не приводит к верным количественным результатам в приложении к атомам с более, чем одним электроном, но зато очень удобна при качественной интерпретации атомных явлений. Точную теорию атома дает квантовая механика.
Дискретная природа микромира. Открытие атомного строения вещества оказалось первым шагом на пути открытий дискретной природы микромира. Не только массы и электрические заряды микротел дискретны, но и динамические величины, описывающие состояния микросистем, такие, как энергия, момент количества движения, также дискретны и характеризуются скачкообразным изменением своих численных значений.
Ядерные структуры
Простейшие синтаксические модели, являющиеся основой речевой деятельности, поскольку они используются для разнообразных трансформаций по требованиям контекста.
Словарь-справочник лингвистических терминов. Изд. 2-е. - М.: Просвещение . Розенталь Д. Э., Теленкова М. А. . 1976 .
Смотреть что такое "ядерные структуры" в других словарях:
ядерные структуры - простейшие синтаксические модели данного языка, являющиеся основой речевой деятельности в том смысле, что пользующиеся данным языком подвергают эти модели разнообразным трансформациям в зависимости от ее требований контекста. Ср. ядерные… …
Превращения ат. ядер при вз ствии с ч цами, в т. ч. с g квантами или друг с другом. Для осуществления Я. р. необходимо сближение ч ц (двух ядер, ядра и нуклона и т. д.) на расстояние 10 13 см. Энергия налетающих положительно заряж. ч ц должна… … Физическая энциклопедия
ядерные фибриллы - Нитевидные внутриядерные структуры, являющиеся фрагментами ядерного скелета [Арефьев В.А., Лисовенко Л.А. Англо русский толковый словарь генетических терминов 1995 407с.] Тематики генетика EN nuclear fibrils … Справочник технического переводчика
Превращения атомных ядер при взаимодействии с элементарными частицами, γ квантами или друг с другом. Для осуществления Я. р. необходимо сближение частиц (двух ядер, ядра и нуклона и т. д.) на расстояние Ядерные реакции 10 13 см. Энергия… …
Обмен веществами между ядром и цитоплазмой клетки осуществляется посредством ядерных пор транспортных каналов, пронизывающих двухслойную ядерную оболочку. Переход молекул из ядра в цитоплазму и в обратном направлении называется ядерно… … Википедия
Сильное взаимодействие (цветовое взаимодействие, ядерное взаимодействие) одно из четырёх фундаментальных взаимодействий в физике. Сильное взаимодействие действует в масштабах атомных ядер и меньше, отвечая за притяжение между нуклонами в ядрах и … Википедия
Nuclear fibrils ядерные фибриллы. Hитевидные внутриядерные структуры, являющиеся фрагментами ядерного скелета
ядерные предложения - простейшие синтаксические построения данного языка, в которых предметы обозначены существительными, процессы глаголами, а признаки прилагательными и наречиями, от которых путем серии трансформаций образуются поверхностные структуры … Толковый переводоведческий словарь
ядерные реакции - превращение атомов ядер при соударении с другими ядрами, элементарными частицами или гамма квантами. При бомбардировке тяжелых ядер более легкими получены все трансурановые элементы. Сокращенно ядерную реакцию, например, типа… … Энциклопедический словарь по металлургии
Ядерные процессы, в которых вносимая в атомное ядро энергия передаётся преимущественно одному или небольшой группе нуклонов (См. Нуклоны). П. я. р. многообразны, они вызываются всевозможными налетающими частицами (от γ квантов до… … Большая советская энциклопедия
Книги
- Инновационная деятельность в атомной отрасли (на примере стратегии развития ядерных топливных циклов, включая инновационные). Книга 1. Основные принципы инновационной политики , А. В. Путилов, А. Г. Воробьев, М. Н. Стриханов. В учебном пособии раскрываются роль и место инноваций в общественном развитии на примере атомной отрасли; цели и задачи национальной инновационной политики. Рассмотрены инструменты…
- Введение в физику микромира. Физика частиц и ядер , Л. И. Сарычева. В настоящей книге представлены основные характеристики фундаментальных и элементарных частиц и процессы, происходящие с ними в различных типах взаимодействий. Описана современная…
В процессе эволюции претерпевали ряд изменений. Появлению новых органелл предшествовали преобразования в атмосфере и литосфере молодой планеты. Одним из значительных приобретений стало клеточное ядро. Эукариотические организмы получили, благодаря наличию обособленных органелл, существенные преимущества перед прокариотами и быстро стали доминировать.
Клеточное ядро, строение и функции которого несколько отличаются в разных тканях и органах, позволило повысить качество биосинтеза РНК и передачу наследственной информации.
Происхождение
На сегодняшний день есть две основные гипотезы об образовании эукариотической клетки. Согласно симбиотической теории органеллы (например, жгутики или митохондрии) когда-то были отдельными прокариотическими организмами. Предки современных эукариот поглотили их. В результате образовался симбиотический организм.
Ядро при этом сформировалось в результате выпячивания внутрь участка цитоплазматической было необходимым приобретением на пути освоения клеткой нового способа питания, фагоцитоза. Захват пищи сопровождался повышением степени подвижности цитоплазмы. Генофоры, представлявшие собой генетический материал прокариотической клетки и прикреплявшиеся к стенкам, попадали в зону сильного «течения» и нуждались в защите. В результате и образовалось глубокое впячивание участка мембраны, содержавшего прикрепленные генофоры. В пользу этой гипотезы свидетельствует тот факт, что оболочка ядра неразрывно связана с цитоплазматической мембраной клетки.
Существует и другая версия развития событий. Согласно вирусной гипотезе происхождения ядра, оно сформировалось в результате заражения клетки древней археи. В нее внедрился ДНК-вирус и постепенно получил полный контроль над жизненными процессами. Ученые, считающие эту теорию более правильной, приводят массу доводов в ее пользу. Однако на сегодняшний день нет исчерпывающего доказательства ни для одной из существующих гипотез.
Одно или несколько
Большая часть клеток современных эукариот имеет ядро. Подавляющее их число содержит только одну подобную органеллу. Существуют, однако, и клетки, которые утратили ядро по причине некоторых функциональных особенностей. К ним относятся, например, эритроциты. Встречаются и клетки с двумя (инфузории) и даже несколькими ядрами.
Структура клеточного ядра

Вне зависимости от особенностей организма, строение ядра характеризуется набором типичных органелл. От внутреннего пространства клетки оно отгорожено двойной мембраной. Внутренние и внешние ее прослойки в некоторых местах сливаются, образуя поры. Их функция заключается в обмене веществ между цитоплазмой и ядром.
Пространство органеллы заполнено кариоплазмой, также называемой ядерным соком или нуклеоплазмой. В ней размещается хроматин и ядрышко. Иногда последний из названных органоид клеточного ядра присутствует не в единственном экземпляре. У некоторых же организмов ядрышки, наоборот, отсутствуют.
Мембрана
Ядерная оболочка образована липидами и состоит из двух слоев: наружного и внутреннего. По сути, это та же клеточная мембрана. Ядро сообщается с каналами эндоплазматической сети через перинуклеарное пространство, полость, образованную двумя слоями оболочки.
Наружная и внутренняя мембрана имеют свои особенности в строении, однако в целом довольно похожи.
Ближайший к цитоплазме
Наружный слой переходит в мембрану эндоплазматической сети. Ее основное отличие от последней — значительно более высокая концентрация белков в структуре. Мембрана, непосредственно контактирующая с цитоплазмой клетки, покрыта слоем рибосом с наружной стороны. С внутренней мембраной она соединяется многочисленными порами, представляющими собой довольно крупные белковые комплексы.
Внутренний слой
Обращенная в клеточное ядро мембрана, в отличие от наружной, гладкая, не покрытая рибосомами. Она ограничивает кариоплазму. Характерная особенность внутренней мембраны — слой ядерной ламины, выстилающий ее со стороны, соприкасающейся с нуклеоплазмой. Эта специфическая белковая структура поддерживает форму оболочки, участвует в регуляции экспрессии генов, а также способствует прикреплению хроматина к мембране ядра.
Обмен веществ
Взаимодействие ядра и цитоплазмы осуществляется через Они представляют собой довольно сложные структуры, образованные 30 белками. Количество пор на одном ядре может быть разным. Он зависит от типа клетки, органа и организма. Так, у человека клеточное ядро может иметь от 3 до 5 тысяч пор, у некоторых лягушек оно доходит до 50 000.

Главная функция пор — обмен веществ между ядром и остальным пространством клетки. Некоторые молекулы проникают сквозь поры пассивно, без дополнительных затрат энергии. Они обладают небольшими размерами. Транспортировка крупных молекул и надмолекулярных комплексов требует расхода определенного количества энергии.
Из кариоплазмы в клетку попадают синтезируемые в ядре молекулы РНК. В обратном направлении транспортируются белки, необходимые для внутриядерных процессов.
Нуклеоплазма
Строение ядерного сока меняется в зависимости от состояния клетки. Их два — стационарное и возникающее в период деления. Первое характерно для интерфазы (время между делениями). При этом ядерный сок отличается равномерным распределением нуклеиновых кислот и неструктурированными молекулами ДНК. В этот период наследственный материал существует в виде хроматина. Деление клеточного ядра сопровождается преобразованием хроматина в хромосомы. В это время изменяется строение кариоплазмы: генетический материал приобретает определенную структуру, ядерная оболочка разрушается, и кариоплазма смешивается с цитоплазмой.
Хромосомы

Основные функции нуклеопротеидных структур преобразованного на время деления хроматина — хранение, реализация и передача наследственной информации, которую содержит клеточное ядро. Хромосомы характеризуются определенной формой: делятся на части или плечи первичной перетяжкой, также называемой целомерой. По ее расположению выделяют три типа хромосом:
- палочкообразные или акроцентрические: для них характерно размещение целомеры практически на конце, одно плечо получается очень маленьким;
- разноплечие или субметацентрические обладают плечами неравной длины;
- равноплечие или метацентрические.
Набор хромосом в клетке называется кариотипом. У каждого вида он фиксирован. При этом разные клетки одного организма могут содержать диплоидный (двойной) или гаплоидный (одинарный) набор. Первый вариант характерен для соматических клеток, в основном составляющих тело. Гаплоидный набор — привилегия половых клеток. Соматические клетки человека содержат 46 хромосом, половые — 23.
Хромосомы диплоидного набора составляют пары. Одинаковые нуклеопротеидные структуры, входящие в пару, называются аллельными. Они имеют одинаковое строение и выполняют одни и те же функции.
Структурной единицей хромосом является ген. Он представляет собой участок молекулы ДНК, кодирующий определенный белок.
Ядрышко

Клеточное ядро обладает еще одним органоидом — это ядрышко. Оно не отделяется от кариоплазмы мембраной, но при этом его легко заметить во время изучения клетки с помощью микроскопа. Некоторые ядра могут иметь несколько ядрышек. Существуют и такие, в которых подобные органоиды отсутствуют совсем.
По форме ядрышко напоминает сферу, имеет достаточно небольшие размеры. В его состав входят различные белки. Основная функция ядрышка — синтез рибосомных РНК и самих рибосом. Они необходимы для создания полипептидных цепей. Ядрышки образуются вокруг специальных участков генома. Они получили название ядрышковых организаторов. Здесь содержатся гены рибосомной РНК. Ядрышко, кроме прочего, является местом с наибольшей концентрацией белка в клетке. Часть белков необходима для выполнения функций органоида.
В составе ядрышка выделяют два компонента: гранулярный и фибриллярный. Первый представляет собой созревающие субъединицы рибосом. В фибриллярном центре осуществляется Гранулярный компонент окружает фибриллярный, расположенный в центре ядрышка.
Клеточное ядро и его функции
Роль, которую играет ядро, неразрывно связана с его строением. Внутренние структуры органоида совместно реализуют важнейшие процессы в клетке. Здесь размещается генетическая информация, которая определяет строение и функции клетки. Ядро отвечает за хранение и передачу наследственной информации, осуществляющееся во время митоза и мейоза. В первом случае дочерняя клетка получает идентичный материнскому набор генов. В результате мейоза образуются половые клетки с гаплоидным набором хромосом.

Другая не менее важная функция ядра — регуляция внутриклеточных процессов. Она осуществляется в результате контроля синтеза белков, отвечающих за строение и функционирование клеточных элементов.
Влияние на белковый синтез имеет еще одно выражение. Ядро, контролируя процессы внутри клетки, объединяет все ее органоиды в единую систему с отлаженным механизмом работы. Сбои в нем приводят, как правило, к гибели клетки.
Наконец, ядро является местом синтеза субъединиц рибосом, которые отвечают за образование все того же белка из аминокислот. Рибосомы незаменимы в процессе транскрипции.

Представляет собой более совершенную структуру, чем прокариотическая. Появление органоидов с собственной мембраной позволило повысить эффективность внутриклеточных процессов. Формирование ядра, окруженного двойной липидной оболочкой, играло в этой эволюции очень важную роль. Защита наследственной информации мембраной позволила освоить древним одноклеточным организмам новые способы жизнедеятельности. Среди них был фагоцитоз, который по одной из версий привел к появлению симбиотического организма, позже ставшего прародителем современной эукариотической клетки со всеми характерными для нее органоидами. Клеточное ядро, строение и функции некоторых новых структур позволили задействовать кислород в метаболизме. Следствием этого стало кардинальное изменение в биосфере Земли, была заложена основа для формирования и развития многоклеточных организмов. Сегодня эукариотические организмы, к которым относится и человек, доминируют на планете, и ничто не предвещает изменений в этом плане.







