Красное ядро
Передние и задние бугры четверохолмия.
Мозжечок.
Белое вещество мозжечка – проводящие пути мозжечка. Среди БВ находятся ядра мозжечка. В мозжечок поступают сигналы от всех структур, связанных с движением. Там они обрабатываются, затем из мозжечка поступает огромный поток тормозных влияний на СМ.
Средний мозг – четверохолмие, черная субстанция, ножки мозга.
Передние бугры – первичная зрительная зона – формируют ориентировочный рефлекс на зрительный сигнал
Задние бугры – первичная слуховая зона – формируют ориентировочный рефлекс на звуковой сигнал
Функция - сторожевые рефлексы (ориентировочные)
Тонус скелетной мускулатуры
Перераспределение тонуса при изменении позы
Упорядочивать взаимоотношение между мышцами сгибателями и разгибателями
Децереберационная ригидность – повреждение красного ядра, резко повышается возбудимость/тонус более сильных мышц
Черная субстанция – источник дофамина
Тормозная функция базальных ганглиев, не дает возбуждать зоны больших полушарий
Тонус скелетных мышц, отвечающих за тонкие инструментальные движения
Пример дисфункции: болезнь Паркинсона
Таламус – поступают сигналы со всех рецепторов кроме обонятельного, его называют коллектором афферентный импульсов.
Прежде чем попасть в кору, информация поступает в таламус. Если таламус разрушен, то кора не получает эту информацию. Если в коленчатые тела (одни из ядер таламуса) поступают зрительные сигналы, то уходят сразу в затылочную долю коры полушарий. Тоже и со слуховой, только она идет в височную. В таламусе обрабатывается информация и выбирается наиболее адекватная
В таламусе десятки ядер, которые делятся на 2 группы: специфические и неспецифические.
Когда поступает информация в специфические ядра таламуса, то в коре возникают вызванные ответы, но ответы возникают в строго выбранных участках полушарий. Информация от неспецифических ядер таламуса поступает ко всей коре больших полушарий. Это происходит, чтобы повысить возбудимость всей коры, чтобы она более четко воспринимала специфическую информацию.
Адекватная боль возникает с участием лобной, теменной коры, таламуса. Таламус - высший центр болевой чувствительности. При разрушении одних ядер таламуса возникает невыносимая боль, при разрушении других ядер полностью теряется болевая чувствительность.
Неспецифические ядра по функции очень похожи на ретикулярную формацию, их еще называют ретикулярными ядрами.
И.И. Сеченов 1864 – открыл ретикулярную формацию, опыты на лягушках. Доказал, что в ЦНС наряду с явлениями возбуждения, есть явления торможения.
Ретикулярная формация – поддерживает кору в состоянии бодрствования. Тормозные влияния на СМ.
Мозолистое тело – плотный пучок нервных волокон, соединяет полушария, обеспечивает их совместную работу.
Гипоталамус – связан с гипофизом. Гипофиз – железа внутренней секреции, главная. В ней вырабатываются тропные гормоны, которые влияют на работу остальных эндокринных желез.
Нейросекреторные клетки гипоталамуса выделяют нейрогормоны:
Статины - тормозят выработку тропных гормонов гипофиза
Либерины – усиливают выработку тропных гормонов гипофиза
Функции - высший цент регуляции эндокринных желез
Нейросекреторные клетки, аксоны которых доходят до гипофиза и выделяют в гипофиз гормоны:
Окситоцин – обеспечивает сокращение матки при родах
Антидиуретический гормон – регулирует работу почек
Клетки гипоталамуса чувствительны к уровню половых гормонов (эстроген и андроген) и в зависимости от того, какие преобладают у человека, возникает та или иная половая мотивация. Клетки гипоталамуса чувствительны к температуре крови, регулирует теплоотдачу.
Главный сигнал голода – уровень глюкозы в крови. Только в гипоталамусе есть глюкорецептивные клетки, чувствительные к уровню глюкозы в крови. Собраны вместе и образуют центр голода.
Центр насыщения – возникновение чувства сытости.
Пример дисфункции: Булимия – заболевания центра сытости
Осморецептивные клетки – чувствительные к уровню солей в крови, возбуждаются – возникает чувство жажды.
На уровне гипоталамуса возникают только мотивации, а для их выполнения нужно включить кору.
Внутри него расположена полость третьего мозгового желудочка. В состав промежуточного мозга входят:
Зрительный мозг
Таламус
Эпиталамус (надталамическая область – эпифиз, поводки, спайка поводков, треугольники поводков)
Метаталамус (заталамическая область – медиальные и латеральные коленчатые тела)
Гипоталамус (подталамическая область)
Передняя гипоталамическая область (зрительная — зрительный перекрест, тракт)
Промежуточная гипоталамическая область (серый бугор, воронка, гипофиз)
Задняя гипоталамическая область (сосочковые тела)
Собственно подталамическая область (заднее гипоталамическое ядро Люизи)
Таламус
Зрительный бугор состоит из серого вещества, разделенного прослойками белого вещества на отдельные ядра. Происходящие их них волокна образуют лучистый венец, связывающий таламус с другими отделами мозга.
Таламус является коллектором всех афферентных (сенсорных) путей, идущих к коре головного мозга. Это ворота на пути к коре, через которые проходит вся информация от рецепторов.
Ядра таламуса:
- Специфические — переключение афферентной импульсации в строго локализованные зоны коры.
1.1. Релейные (переключательные)
1.1.1. Сенсорные (вентральные задние, вентральное промежуточное ядро) переключение афферентной импульсации в сенсорные зоны коры .
1.1.2. Несенсорные – переключение несенсорной информации в кору.
- Лимбические ядра (передние ядра) – подкорковый центр обоняния. Передние ядра таламуса — лимбическая кора -гиппокамп-гипоталамус-мамиллярные тела гипоталамуса — передние ядра таламуса (круг ревербации Пейпеца — формирование эмоций).
- Моторные ядра : (вентральные) переключают импульсы от базальных ганглиев, зубчатого ядра мозжечка, красного ядра в моторную и премоторную зону КГМ (передача сложных двигательных программ, образованных в мозжечке и базальных ганглиях).
1.2. Ассоциативные (интегративная функция, получают информацию от других ядер таламуса, посылают импульсы в ассоциативные участки КГМ , существует обратная связь)
1.2.1. Ядра подушки — импульсы от коленчатых тел и неспецифических ядер таламуса, в височно-теменно-затылочные зоны КГМ, участвующие в гностических, речевых и зрительных реакциях (интеграция слова со зрательным образом), восприятии схемы тела. Электростимуляция подушки ведёт к нарушению называния предметов, разрушение подушки – нарушение схемы тела, устраняет тяжёлые боли.
1.2.2. Медиодорсальное ядро – от гипоталамуса, миндалины, гиппокампа, таламических ядер, центрального серого вещества ствола, к ассоциативной лобной и лимбической коре. Формирование эмоций и поведенческой двигательной активности, участие в механизмах памяти. Разрушение – устраняет страх, тревогу, напряжённость, страдание от боли, но снижается инициатива, безразличие, гипокинезия.
1.2.3. Латеральные ядра – от коленчатых тел, вентрального ядра таламуса, к теменной коре (гнозис, праксис, схема тела.)
- Неспецифические ядра – (интраламинарные ядра, ретикулярное ядро) передача сигналов во все участки КГМ . Множество входящих и выходящих волокон, аналог РФ ствола – интегрирующая роль между стволом мозга, мозжечком и базальными ганглиями, новой и лимбической корой. Модулирующее влияние, обеспечивают тонкое регулирование поведения, «плавную настройку» ВНД.
Метаталамус Медиальные коленчатые тела вместе с нижними бугорками четверохолмия среднего мозга образуют подкорковый центр слуха. Они играют роль коммутационных центров для нервных импульсов, направляющихся в кору головного мозга. На нейронах ядра медиального коленчатого тела заканчиваются волокна латеральной петли. Латеральные коленчатые тела вместе с верхними бугорками четверохолмия и подушкой зрительного бугра являются подкорковыми центрами зрения. Они представляют собой коммуникационные центры, на которых заканчивается зрительный тракт, и в которых прерываются пути, проводящие нервные импульсы к зрительным центрам коры больших полушарий.
Эпиталамус Эпифиз связывают с теменным органом некоторых высших рыб и пресмыкающихся. У круглоротых он сохранил до известной степени строение глаза, у бесхвостых земноводных находится в редуцированном виде под кожей головы. У млекопитающих и человека Эпифиз имеет железистое строение и является железой внутренней секреции (гормон – мелатонин).
Эпифиз (шишковидная железа) относится к железам внутренней секреции. Он вырабатывает серотонин, из которого затем образуется мелатонин. Последний является антагонистом меланоцитостимулирующего гормона гипофиза, а также половых гормонов. Деятельность эпифиза зависит от освещенности, т.е. проявляется циркадность ритма, а это регулирует репродуктивную функцию организма.
Гипоталамус
Гипоталамическая область содержит сорок две пары ядер, которые делятся на четыре группы: переднюю, промежуточную, заднюю и дорсолатеральную.
Гипоталамус это вентральная часть промежуточного мозга, анатомически состоит из преоптическая области, области перекреста зрительных нервов, серого бугра и воронки, сосцевидных тел. Выделяют следующие группы ядер:
- Передняя группа ядер (кпереди от серого ядра) – преоптические ядра, супрахиазматическое, супраоптическое, паравентрикулярное
- Промежуточная (туберальная) группа (в области серого бугра и воронки) – дорсомедиальное, вентромедиальное, аркуатное (инфундибулярное), дорсальное подбугорное, заднее ПВЯ и собственные ядра бугра и воронки. Первые две группы ядер являются нейросекреторными.
- Задняя – ядра сосочковых тел (подкорковый центр обоняния)
- Субталамическое ядро Луиса (интреграционная функция
В гипоталамусе имеется самая мощная в мозге сеть капилляров и самый большой уровень локального кровотока до 2900 капилляров на мм квадратный). Высока проницаемость капилляров, т.к. гипоталамус имеет клетки избирательно чувствительные к изменениям параметров крови: изменениям рН, содержанию ионов калия, натрия, напряжению кислорода, углекислого газа. Супраоптическое ядро имеет осморецепторы , вентромедиальное ядро имеет хеморецепторы , чувствительные к уровню глюкозы, в переднем гипоталамусе рецепторы к половым гормонам . Есть терморецепторы . Чувствительные нейроны гипоталамуса не адаптируются, и находятся возбуждении до тех пор, пока та или иная константа в организме не нормализуется. Эфферентные влияния гипоталамус осуществляет с помощью симпатической и парасимпатической нервных систем, и эндокринных желёз. Здесь расположены центры регуляции различных видов обменов: белкового, углеводного, жирового, минерального, водного, а также центры голода, жажды, насыщения, удовольствия. Гипоталамическую область относят к высшим подкорковым центрам вегетативной регуляции. Вместе с гипофизом она образует гипоталамо-гипофизарную систему, посредством которой в организме сопрягается нервное и гормональное регулирование.
В гипоталамической области синтезируются эндорфины и энкефалины, входящие в состав естественной противоболевой системы и влияющих на психику человека.
Нервные пути к гипоталамусу идут от лимбической системы, КГМ, базальных ганглиев, РФ ствола. От гипоталамуса – в РФ, моторные и вегетативные центры ствола вегетативные центры спинного мозга, от мамиллярных тел к передним ядрам таламуса, далее в лимбическую систему, от СОЯ и ПВЯ к нейрогипофизу, от вентромедиального и инфундибулярного – к аденогипофизу, также имеются связи с лобной корой и полосатым телом.
Гормоны СОЯ и ПВЯ:
- АДГ (вазопрессин)
- Окситоцин
Гормоны медиобазального гипоталамуса: вентромедиального и инфундибулярного ядер:
Либерины (рилизинги) кортиколиберин, тиролиберин, люлиберин, фоллилиберин, соматолиберин, пролактолиберин, меланолиберин
Статины (ингибины) соматостатин, пролактостатин и меланостатин
Функции:
- Поддержание гомеостаза
- Интегративный центр вегетативных функций
- Высший эндокринный центр
- Регуляция теплового баланса (передние ядра — центр теплоотдачи, задние — центр теплообразования)
- Регулятор цикла «сон-бодрствование» и других биоритмов
- Роль в пищевом поведении (средняя группа ядер: латеральное ядро — центр голода и вентромедиальное ядро – центр насыщения)
- Роль в половом, агрессивно-оборонительном поведении. Раздражение передних ядер стимулирует половое поведение, раздражение задних ядер угнетает половое развитие.
- Центр регуляции различных видов обменов: белкового, углеводного, жирового, минерального, водного.
- Является элементом антиноцицептивной системы (центр удовольствия)
Таламус является частью промежуточного мозга. Это структура, в которой происходит обработка и интеграция практически всех сигналов, идущих в кору большого мозга от спинного, среднего мозга, мозжечка, базальных ганглиев головного мозга. Зрительный бугор в целом является подкорковой «станцией» для всех видов чувствительности. Здесь раздражения внешней и внутренней среды интегрируются, после чего поступают в кору большого мозга (коммутатор мозга).
Ядра таламуса функционально по характеру входящих и выходящих из них путей делятся на следующие группы.
- 1. Специфические ядра. К ним относятся переднее вентральное, медиальное, вентролатеральиое, постлатеральное, постмедиальное, латеральное и медиальное коленчатые тела. Основной функциональной единицей специфических таламических ядер являются «релейные» нейроны, у которых мало дендритов и длинный аксон; их функция заключается в переключении информации, идущей в кору большого мозга от кожных, мышечных и других рецепторов. От специфических ядер информация о характере сенсорных стимулов поступает в строго определенные участки III-IV слоев коры большого мозга (соматотопическая локализация). Нарушение функции специфических ядер приводит к выпадению конкретных видов чувствительности. Вентролатеральное ядро передает сигналы к двигательной коре, в связи с чем, является мишенью при стереотаксических операциях у больных с дискинезиями (болезнь Паркинсона, спастическая кривошея и т.д.).
- 2. Ассоциативные ядра таламуса представлены передним медио-дорсальным, латеральным дорсальным ядрами и подушкой. Переднее ядро связано с лимбической корой (поясной извилиной), медиодорсальное - с лобной долей коры, латеральное дорсальное - с теменной, полушка - с ассоциативными зонами теменной и височной долями коры большого мозга. Основными клеточными структурами этих ядер являются мультиполярные нейроны, способные выполнять полисенсорные функции.
- 3. Неспецифические ядра таламуса представлены срединным центром, парацентральным ядром, центральным медиальным и латеральным, субмедиальным, вентральным передним, парафасцикулярным комплексами, ретикулярным ядром, перивентрикулярной и центральной серой массой. Нейроны этих ядер образуют свои связи по ретикулярному типу. Их аксоны поднимаются в кору большого мозга и контактируют со всеми ее слоями, образуя не локальные, а диффузные связи. К неспецифическим ядрам поступают связи из РФ ствола мозга, гипоталамуса, лимбической системы, базальных ганглиев, специфических ядер таламуса. Возбуждение неспецифических ядер вызывает генерацию в коре специфической веретенообразной электрической активности, свидетельствующей о развитии сонного состояния.
Сложное строение таламуса, наличие в нем взаимосвязанных специфических, неспецифических и ассоциативных ядер, позволяет ему организовывать такие двигательные реакции, как сосание, жевание, глотание, смех. Двигательные реакции интегрируются в таламусе с вегетативными процессами, обеспечивающими эти движения.
Таламус (зрительные бугры)
Нейроны таламуса образуют 40 ядер. Топографически ядра таламуса подразделяются на передние, срединные и задние. Функционально эти ядра можно разделить на две группы: специфические и неспецифические.
Специфические ядра входят в состав специфических проводящих путей. Это восходящие пути, которые передают информацию от рецепторов органов чувств к проекционным зонам коры полушарий большого мозга.
Важнейшими из специфических ядер являются латеральное коленчатое тело, участвующее в передаче сигналов от фоторецепторов и медиальное коленчатое тело, передающее сигналы от слуховых рецепторов.
Неспецифические яра таламуса относят к ретикулярной формации. Они выполняют роль интегративных центров и оказывают преимущественно активирующее восходящее влияние на кору полушарий большого мозга:
1 — передняя группа (обонятельные); 2 — задняя группа (зрительные); 3 — латеральная группа (общая чувствительность); 4 — медиальная группа (экстрапирамидная система; 5 — центральная группа (ретикулярная формация).

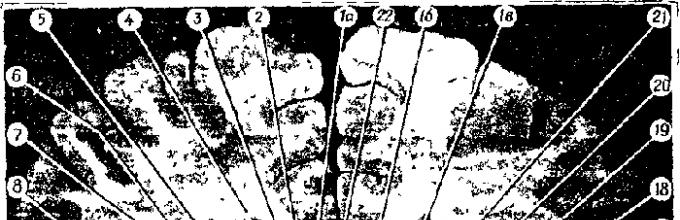
Фронтальный срез головного мозга на уровне середины зрительного бугра. 1а — переднее ядро зрительного бугра. 16 — медиальное ядро зрительного бугра, 1в — латеральное ядро зрительного бугра, 2 — боковой желудочек, 3 — свод, 4 — хвостатое ядро, 5 — внутренняя капсула, 6 — наружная капсула, 7 — внешняя капсула (capsula extrema), 8 — вентральное ядро зрительного бугра, 9 — субталамическое ядро, 10 — третий желудочек, 11 — ножка мозга. 12 — мост, 13 — межножковая ямка, 14 — ножка гиппокампа, 15 — нижний рог бокового желудочка. 16 — черное вещество, 17 — островок. 18 — бледный шар, 19 — скорлупа, 20 — поля Фореля Н; и ЬЬ. 21 — межталамическое сращение, 22 — мозолистое тело, 23 — хвост хвостатого ядра.
Активацию нейронов неспецифических ядер таламуса особенно эффективно вызывают болевые сигналы (таламус – высший центр болевой чувствительности).
Повреждения неспецифических ядер таламуса приводят также к нарушению сознания: потерей активной связи организма с окружающей средой.
Подбугорье (гипоталамус)
Гипоталамус образован группой ядер, ррасположенных у основания мозга. Ядра гипоталамуса представляют собой подкорковые центры автономной нервной системы всех жизненно важных функций организма.
Топографически гипоталамус разделяется на преоптическую область, области переднего, среднего и заднего гипоталамуса.
Studepedia.org — это Лекции, Методички, и много других полезных для учебы материалов
Все ядра гипоталамуса парные.

Метаталамус и гипоталамус. 1 — водопровод 2 — красное ядро 3 — покрышка 4 — черная субстанция 5 — ножка мозга 6 -сосцевидные тела 7 — переднее продырявленное вещество 8 — обобнятельный треугольник 9 — воронка 10 — зрительный перекрест 11. зрительный нерв 12 — серый бугор 13-заднее продырявленное вещество 14 — наружное коленчатое тело 15 — медиальное коленчатое тело 16 — подушка 17 — зрительный тракт

Подбугорная область (hypothalamus)
а — вид снизу; б — серединное сагиттальное сечение.
Зрительная часть (pars optica): 1 — конечная пластинка; 2 — зрительный перекрест; 3 — зрительный тракт; 4 — серый бугор; 5 — воронка; 6 — гипофиз;
Обонятельная часть: 7 — сосковидные тела — подкорковые обонятельные центры; 8 — подбугорная область в узком смысле слова является продолжением ножек мозга, содержит черное вещество, красное ядро и Люисово тело, которое является звеном экстрапирамидной системы и вегетативным центром; 9 — подбугорная Монроева борозда; 10 — турецкое седло, в ямке которого находится гипофиз.

Главные ядра гипоталамуса

Схема нейросекреторных ядер подбугорной области (Hypothalamus). 1 — nucleus supraopticus; 2 — nucleus preopticus; 3 — nuclius paraventricularis; 4 — nucleus infundibularus; 5 — nucleus coгрoris mamillaris; 6 — зрительный перекрест; 7 — гипофиз; 8 — серый бугор; 9 — сосцевидное тело; 10 мост.
В преоптическую область входят перивентрикулярное, медиальное и латеральное преоптические ядра.
В группу переднего гипоталамуса относят супраоптическое, супрахиазматическое и паравентрикулярное ядра.
Средний гипоталамус составляет вентромедиальное и дорсомедиальное ядра.
В заднем гипоталамусе различают заднее гипоталамическое, перифорникальное и мамиллярное ядра.
Связи гипоталамуса обширны и сложны. Афферентные сигналы в гипоталамус поступают от коры больших полушарий, подкорковых ядер и от таламуса. Основные эфферентные пути доходят до среднего мозга, таламуса и подкорковых ядер.
Гипоталамус является высшим центром регуляции сердечно-сосудистой системы, водно-солевого, белкового, жирового, углеводного обменов. В этой области мозга расположены центры, связанные с регуляцией пищевого поведения. Важная роль гипоталамуса – регуляция. Электрическое раздражение задних ядер гипоталамуса приводит к гипертермии, в результате повышения обмена веществ.
Гипоталамус принимает также участие в поддержании биоритма “сон-бодрствование”.
Ядра переднего гипоталамуса связаны с гипофизом и осуществляют транспорт биологически активных веществ, которые вырабатываются нейронами этих ядер. Нейроны преоптического ядра вырабатывают рилизинг-факторы (статины и либерины), контролирующие синтез и высвобождение гормонов гипофиза.
Нейроны преоптического, супраоптического, паравентрикулярного ядер вырабатывают истинные гормоны – вазопрессин и окситоцин, которые по аксонам нейронов спускаются в нейрогипофиз, где хранятся до высвобождения – поступления в кровь.
Нейроны передней доли гипофиза вырабатывают 4 вида гормонов: 1) соматотропный гормон, регулирующий рост; 2) гонадотропный гормон, способствующий росту половых клеток, желтого тела, усиливает выработку молока; 3) тиреотропный гормон – стимулирует функцию щитовидной железы; 4) адренокортикотропный гормон – усиливает синтез гормонов коры надпочечников.
Промежуточная доля гипофиза выделяет гормон интермедин, влияющий на пигментацию кожи.
Задняя доля гипофиза выделяет два гормона – вазопрессин, влияющий на гладкую мускулатуру артериол, и окситоцин – действует на гладкую мускулатуру матки и стимулирует выделение молока.
Гипоталамус играет также важную роль в эмоциональном и половом поведении.
В состав эпиталамуса (шишковидная железа) входит эпифиз. Гормон эпифиза – мелатонин – тормозит в гипофизе образование гонадотропных гормонов, а это в свою очередь задерживает половое развитие.
Неспецифическое ядро
Cтраница 1
Неспецифические ядра являются по своему происхождению более древними и включают срединные и интраламинарные ядра, а также медиальную часть переднего вентрального ядра. Нейроны неспецифических ядер сначала передают сигналы в подкорковые структуры, от которых импульсация поступает параллельно в разные отделы коры. Неспецифические ядра являются продолжением ретикулярной формации среднего мозга, представляя собой ретикулярную формацию таламуса.
Функции Промежуточного мозга
Электрическое раздражение неспецифических ядер таламуса вызывает в коре больших полушарий периодические колебания потенциалов, синхронные с ритмом активности таламических структур. Реакция в коре возникает с большим скрытым периодом и значительно усиливается при повторении. Таким образом, нейроны коры больших полушарий вовлекаются в процесс активности как бы постепенно. Такая реакция вовлечения коры мозга отличается от специфических ее ответов своей генерализованностью, охватом обширных областей коры. Импульсы, идущие по путям болевой чувствительности, формируются при раздражении различных областей тела и внутренних органов. Скрытые периоды ответов в таламусе отличаются большой длительностью и вариабельностью.
Другой тип окончаний таламокорти-кальных проекций образуется аксонами нейронов неспецифических ядер таламуса.
При регистрации электрической активности различных отделов головного мозга кролика было обнаружено, что реакции в виде увеличения числа мыленных волн и веретен возникают одновременно во всех отведениях (при скорости записи 15 мм / с), а наиболее интенсивную реакцию наблюдали в гипоталамусе, затем следовала кора сензомоторная, зрительная, специфические ядра таламу-са, неспецифические ядра таламуса. Можно заключить, что наиболее реактивными образованиями ЦНС при воздействии ПМП являются кора и гипоталамус.
Через неспецифические ядра таламуса в кору мозга поступают восходящие активирующие влияния от ретикулярной формации мозгового ствола. Система неспецифических ядер таламуса осуществляет контроль ритмической активности коры больших полушарий и выполняет функции внутриталамической интегрирующей системы.
Для изучения механизма образования условных рефлексов существенное значение имеет не только точная регистрация самой ответной реакции (слюноотделения, движения и т.п.), но также исследование электрической активности, возникающей в различных мозговых структурах во время действия условного и безусловного раздражителей. Для регистрации электрической активности применяют электроды, хронически вживляемые в различные области или слои коры большого мозга, а также в специфические и неспецифические ядра таламуса, ретикулярную формацию, гиппокамп и другие отделы головного мозга. В опытах с условными рефлексами широко используются микроэлектродные методы, позволяющие регистрировать электрическую активность отдельных нейронов, участвующих в осуществлении условнорефлекторной реакции. Для автоматического анализа электроэнцефалограмм, регистрируемых с различных областей коры, в опытах на животных непосредственно во время условнорефлекторных реакций используют электронно-вычислительные машины.
Неспецифические ядра являются по своему происхождению более древними и включают срединные и интраламинарные ядра, а также медиальную часть переднего вентрального ядра. Нейроны неспецифических ядер сначала передают сигналы в подкорковые структуры, от которых импульсация поступает параллельно в разные отделы коры. Неспецифические ядра являются продолжением ретикулярной формации среднего мозга, представляя собой ретикулярную формацию таламуса.
Нейроны специфического комплекса ядер посылают по направлению к коре аксоны, почти не имеющие коллатералей. В отличие от него нейроны неспецифической системы посылают аксоны, дающие множество коллатералей. Вместе с тем и волокна, приходящие из коры к нейронам специфических ядер, характеризуются топографической локали-зованностью своих окончаний в противоположность широко разветвленной системе диф-фузно оканчивающихся волокон в неспецифических ядрах.
Спиноталамический путь значительно отличается от лемнискового. Его первые нейроны также расположены в спинальном ганглии, откуда они посылают в спинной мозг медленно проводящие немиелинизированные нервные волокна. Эти нейроны имеют большие рецептивные поля, иногда включающие значительную часть кожной поверхности. Вторые нейроны данного пути локализуются в сером веществе спинного мозга, а их аксоны в составе восходящего спиноталамического тракта направляются после перекреста на спинальном уровне в вентробазальный ядерный комплекс таламуса (дифференцированные проекции), а также в вентральные неспецифические ядра таламуса, внутреннее коленчатое тело, ядра ствола мозга и гипоталамус. Локализованные в этих ядрах третьи нейроны спиноталамического пути лишь частично дают проекции в соматосенсорную зону коры.
Страницы: 1
8. Строение и функциональная роль таламуса и гипоталамуса
Таламус (лат. Thalamus, латинское произношение: талямус; от греч. θάλαμος - «бугор») - область головного мозга, отвечающая за перераспределение информации от органов чувств, за исключением обоняния, к коре головного мозга.
Эта информация (импульсы) поступает в ядра таламуса. Сами ядра состоят из серого вещества, которое образовано нейронами. Каждое ядро представляет собой скопление нейронов. Ядра разделяет белое вещество. В таламусе можно выделить четыре основных ядра: группа нейронов перераспределяющая зрительную информацию; ядро перераспределяющее слуховую информацию; ядро перераспределяющее тактильную информацию и ядро перераспределяющее чувство равновесия и баланса. После того как информация о каком-либо ощущении поступила в ядро таламуса, там происходит её первичная обработка, то есть впервые осознается температура, зрительный образ и т. д. Считается, что таламус играет важную роль в осуществлении процессов запоминания. Фиксация информации осуществляется следующим образом: первая стадия формирования энграммы происходит в СС. Это начинается, когда стимул возбуждает периферические рецепторы. От них по проводящим путям нервные импульсы идут в таламус, а затем в корковый отдел. В нем осуществляется высший синтез ощущения. Повреждение таламуса может привести к антероградной амнезии, а также вызвать тремор - непроизвольную дрожь конечностей в состоянии покоя, - хотя эти симптомы отсутствуют, когда пациент выполняет движения осознанно. С таламусом связано редкое заболевание, называемое «фатальная семейная бессонница» . http://www.bibliotekar.ru/447/52.htm medbiol.ru/medbiol/mozg/0001b9d3.htm
Таламус (зрительный бугор, thalamus): общие сведения
Таламус — участок переднего мозга.
Анатомически таламус (зрительный бугор) — парный орган, образованный главным образом серым веществом. Он является подкорковым центром всех видов чувствительности, в нем насчитывают несколько десятков ядер, которые получают информацию от всех органов чувств и передают ее в кору головного мозга. Таламус связан с лимбической системой, ретикулярной формацией, гипоталамусом, мозжечком, базальными ганглиями. Таламус представляет собой яйцевидную массу серого вещества с задним более утолщенным концом (рис. 38 , рис. 39).
Как было уже сказано, таламус — парное образование: существует дорсальный таламус и вентральный таламус.. Между таламусами находится полость III желудочка. Поверхность таламуса, обращенная в полость III желудочка, покрыта тонким слоем серого вещества. Медиальные поверхности правого и левого таламусов соединены между собой межталамическим сращением, лежащим почти посередине. Медиальная поверхность таламуса отделена от верхней тонкой мозговой полоской. Верхняя часть зрительных бугров свободна и обращена в полость центральной части латеральных желудочков. В переднем отделе таламус суживается и заканчивается передним бугорком. Задний конец таламуса утолщен и называется подушкой таламуса. Название "подушка" возникло в связи с тем, что на таламусах лежат полушария конечного мозга, и они покоятся на утолщениях, напоминающих подушку. Латеральная поверхность таламуса прилежит к внутренней капсуле и граничит с хвостатым ядром конечного мозга. Нижняя поверхность таламуса располагается над ножкой мозга, срастаясь с покрышкой среднего мозга.
Прослеживается выраженная эволюционная закономерность изменения количественных соотношений между дорсальным и вентральным таламусом. В процессе эволюции размеры вентральной части таламуса уменьшаются, а дорсальной — увеличиваются. У низших позвоночных развит вентральный таламус, а у млекопитающих преобладают ядра дорсального таламуса. Это обусловлено тем, что дорсальная часть таламуса связана преимущественно с развитием восходящих путей от зрительной системы, слуховой системы и сенсомоторных систем к коре полушарий конечного мозга.
В таламусе оканчиваются аксоны большинства сенсорных нейронов, несущих импульсы в кору головного мозга. Здесь анализируется характер и происхождение этих импульсов, и они передаются в соответствующие сенсорные зоны коры по волокнам, берущим начало в таламусе. Таким образом, таламус играет роль перерабатывающего, интегрирующего и переключающего центра для всей сенсорной информации. Кроме того, в таламусе модифицируется информация, поступающая из определенных зон коры, и полагают, что он участвует в ощущении боли и ощущении удовольствия. В таламусе начинается та область ретикулярной формации, которая имееет отношение к регуляции двигательной активности. Дорсальный участок, лежащий непосредственно перед таламусом — переднее сосудистое сплетение — ответственен за транспорт веществ между спинномозговой жидкостью, находящейся в третьем желудочке, и жидкостью, заполняющей подпаутинное пространство. Таким образом, таламус фильтрует информацию, поступающую от всех рецепторов, осуществляет ее предварительную обработку и после этого направляет ее в различные области коры. Кроме того, таламус осуществляет связи между корой, с одной стороны, и мозжечком и базальными ганглиями с другой.
Иными словами, через таламус сознание контролирует автоматические движения.
Аксоны заднестолбового медиального лемнискового пути и спиноталамического тракта оканчиваются синапсами на нейронах ВПЛ-ядра таламуса. В этом ядре также оканчиваются несколько других параллельных восходящих сенсорных трактов, таких как спиноцервикальный тракт и путь через ядро z . Тройнично-таламические пути от главного сенсорного ядра тройничного нерва и спинального ядра тройничного нерва образуют синапсы в таламическом ВПМ-ядре.
Ответы многих нейронов ВПЛ- иВПМ-ядер аналогичны реакциям нейронов первого и второго порядков восходящих трактов. Среди этих ответов иногда преобладают реакции сенсорных рецепторов определенного типа, причем их рецептивные поля могут быть невелики, хотя обычно обширнее, чем у первичных афферентов.
Эти поля располагаются контрлатерально по отношению к таламическим нейронам, локализация которых топографически связана с местонахождением рецептивных полей, т.е. ВПЛ- и ВПМ-ядер, и имеют соматотопическую организацию. Нижняя конечность представлена нейронами латеральной части ВПЛ-ядра, верхння — нейронами медиальной части ВПЛ-ядра, а лицо — нейронами ВПМ-ядра (рис. 34.10).
Во многих таламических нейронах находятся не только возбуждающие, но и тормозные рецептивные поля. Процесс торможения может реализовываться в ядрах заднего столба или заднем роге спинного мозга, однако тормозные нейронные цепи есть и в таламусе. В ВПЛ- и ВПМ-ядрах присутствуют тормозные интернейроны (у приматов, но не у грызунов), кроме того, проецируются некоторые тормозные интернейроны ретикулярного ядра таламуса. В собственных тормозных нейронах этих ядер и нейронах ретикулярного ядра тормозным медиатором является GABA .
Нейроны ВПЛ- и ВПМ-ядер обладают интересной особенностью: в отличие от активности сенсорных нейронов более низких уровней соматосенсорной системы возбудимость таламических нейронов зависит от стадии цикла "сон-бодрствование" и меняется при анестезии.
Во время дремоты или барбитуратной анестезии таламические нейроны проявляют тенденцию к индукции попеременных последовательностей возбуждающих и тормозных постсинаптических потенциалов. Перемежающиеся разряды, в свою очередь, вызывают периодическую активность нейронов коры мозга. На энцефалограмме это находит отражение в альфа-ритме или залпах веретен. Такое чередование серий возбуждающих и тормозных постсинаптических потенциалов, возможно, отражает уровень возбуждения таламических нейронов, которое опосредуется взаимодействием возбуждающих нейромедиаторных аминокислот с постсинаптическими мембранными рецепторами не NMDA -типа и NMDA-типа. Кроме того, в этом периодическом процессе может участвовать торможение таламических нейронов, опосредуемое возвратными путями ретикулярного ядра.
Спиноталамический тракт и часть тройнично-таламического пути, начинающаяся от спинального ядра тройничного нерва, посылают проекции к центральному латеральному ядру внутрипластинчатого комплекса таламуса. Внутрипластинчатые ядра не имеют соматотопической организации и диффузно проецируются в коре большого мозга, а также в базальных ганглиях. Возможно, проекции центрального латерального ядра в корковой зоне SI участвуют в формировании в этой области реакции пробуждения и механизме избирательного внимания.
После разрушения ВПЛ- и ВПМ-ядер снижается чувствительность контралатеральной стороны туловища и лица. Дефицит касается главным образом сенсорных категорий, связанных с передачей информации по заднестолбовому медиальному лемнисковому тракту и эквивалентной ему системе тройничного нерва. Утрачивается и сенсорно-дкскриминативный компонент болевой чувствительности, но при интактном медиальном таламусе сохраняется мотивационпо-аффективныи компонент, предположительно, благодаря медиальным спиноталамическим и спиноретикулоталамическим проекциям.
У некоторых людей после повреждения соматосенсорного таламуса наступает синдром центральной боли, называемой таламической. Однако боль, не отличающаяся от таламической, может развиться и после повреждении ствола или коры мозга.
См. также рис. 1 , рис.
Промежуточный мозг. Таламус. Ядра таламуса. Гипоталамус. Гормоны СОЯ и ПВЯ.
33 , рис. 42 , рис. 43 , рис. 44 , рис. 59 , рис. 63 , рис. 64 , рис. 75 .
Введение
Таламус (зрительный бугор)
Гипоталамус
Заключение
Медиальное коленчатое тело находится позади подушки таламуса; вместе с нижними холмиками пластинки крыши среднего мозга оно является подкорковым центром слухового анализатора.
Латеральное коленчатое тело располагается книзу от подушки таламуса. Вместе с верхними бугорками четверохолмия оно образует подкорковый центр зрительного анализатора.
Эпиталамус (надталамическая область) включает шишковидное тело (эпифиз), поводки и треугольники поводков . В треугольниках поводков залегают ядра, относящиеся к обонятельному анализатору. Поводки отходят от треугольников поводков, идут каудально, соединяются посредством спайки и переходят в шишковидное тело. Последнее как бы подвешено на них и располагается между верхними бугорками четверохолмия. Шишковидное тело является железой внутренней секреции. Его функции полностью не установлены, предполагается, что оно регулирует наступление полового созревания.
Таламус (зрительный бугор)
Общее строение и расположение таламуса.
Рисунок 1. Промежуточный мозг на сагиттальном разрезе.
Толща серого вещества таламуса разделена вертикальной Y-образной прослойкой (пластинкой) белого вещества на три части - переднюю, медиальную и латеральную.
Медиальная поверхность таламуса хорошо видна на сагиттальном (сагиттальный - стреловидный (лат. "sagitta" - стрела), делящей на симметричные правую и левую половины) разрезе мозга (рис.1). Медиальная (т.е. располагающаяся ближе к середине) поверхность правого и левого таламусов, обращенные друг к другу, образуют боковые стенки III мозгового желудочка (полость промежуточного мозга) посередине они соединены между собой межталамическим сращением .
Передняя (нижняя) поверхность таламусов сращена с гипоталамусом, через нее с каудальной стороны (т.е. находящейся ближе к нижней части тела) в промежуточный мозг входят проводящие пути из ножек мозга.
Латеральная ( т.е. боковая) поверхность таламуса граничит с внутренней капсулой - слоем белого вещества полушарий головного мозга, состоящего из проекционных волокон, соединяющих кору полушарий с нижележащими мозговыми структурами.
В каждой из этих частей таламуса находится несколько групп таламических ядер . Всего в таламусе содержится от 40 до 150 специализированных ядер .
Функциональное значение ядер таламуса.
По топографии ядра таламуса объединяют в 8 основных групп:
1. переднюю группу;
2. медиодорсальную группу;
3. группу ядер средней линии;
4. дорсолатеральную группу;
5. вентролатеральную группу;
6. вентральную заднемедиальную группу;
7. заднюю группу (ядра подушки таламуса);
8. интраламинарную группу.
Ядра таламуса делят на сенсорные ( специфические и неспецифические), моторные и ассоциативные . Рассмотрим основные группы ядер таламуса, необходимые для понимания его функциональной роли в передаче сенсорной информации в кору больших полушарий.
В передней части таламуса располагается передняя группа таламических ядер ( рис.2). Наиболее крупные из них - передневентральное ядро и переднемедиальное ядро. Они получают афферентные волокна от сосцевидных тел - обонятельного центра промежуточного мозга. Эфферентные волокна (нисходящие, т.е. выносящие импульсы из мозга) от передних ядер направляются к поясной извилине коры больших полушарий.
Передняя группа таламических ядер и связанные с нею структуры являются важным компонентом лимбической системы мозга, управляющей психоэмоциональным поведением .

Рис. 2. Топография ядер таламуса
В медиальной части таламуса различают медиодорсалъное ядро и группу ядер средней линии.
Медиодорсальное ядро имеет двусторонние связи с обонятельной корой лобной доли и поясной извилиной больших полушарий, миндалевидным телом и переднемедиальным ядром таламуса. Функционально оно тесно связано также с лимбической системой и имеет двусторонние связи с корой теменной, височной и островковой долей мозга.
Медиодорсальное ядро участвует в реализации высших психических процессов. Его разрушение приводит к снижению беспокойства, тревожности, напряженности, агрессивности, устранению навязчивых мыслей.
Ядра средней линии многочисленны и занимают наиболее медиальное положение в таламусе. Они получают афферентные (т.е. восходящие) волокна от гипоталамуса, от ядер шва, голубого пятна ретикулярной формации ствола мозга и частично от спинно-таламических путей в составе медиальной петли. Эфферентные волокна от ядер средней линии направляются к гиппокампу, миндалевидному телу и поясной извилине больших полушарий, входящих в состав лимбической системы. Связи с корой больших полушарий двусторонние.
Ядра средней линии играют важную роль в процессах пробуждения и активации коры больших полушарий, а также в обеспечении процессов памяти.
В латеральной (т.е. боковой) части таламуса располагаются дорсолатералъная, вентролатеральная, вентральная заднемедиальная и задняя группы ядер.
Ядра дорсолатералъной группы относительно мало изучены. Известно, что они причастны к системе восприятия боли.
Ядра вентролатералъной группы анатомически и функционально различаются между собой. Задние ядра вентролатеральной группы часто рассматриваются как одно вентролатеральное ядро таламуса. Эта группа получает волокна восходящего пути общей чувствительности в составе медиальной петли. Сюда приходят также волокна вкусовой чувствительности и волокна от вестибулярных ядер. Эфферентные волокна, начинающиеся от ядер вентролатеральной группы, направляются в кору теменной доли больших полушарий, куда проводят соматосенсорную информацию от всего тела.
К ядрам задней группы (ядра подушки таламуса) идут афферентные волокна от верхних холмиков четверохолмия и волокна в составе зрительных трактов. Эфферентные волокна широко распространяются в коре лобной, теменной, затылочной, височной и лимбической долей больших полушарий.
Ядерные центры подушки таламуса причастны к комплексному анализу различных сенсорных раздражителей. Они играют значительную роль в перцептивной (связанной с восприятием) и когнитивной (познавательной, мыслительной) деятельности мозга, а также в процессах памяти - хранения и воспроизведения информации.
Интраламинарная группа ядер таламуса лежит в толще вертикальной Y-образной прослойки белого вещества. Интраламинарные ядра взаимосвязаны с базальными ядрами, зубчатым ядром мозжечка и корой больших полушарий.
Эти ядра играют важную роль в активационной системе мозга. Повреждение интраламинарных ядер в обоих таламусах приводит к резкому снижению двигательной активности, а также апатии и разрушению мотивационной структуры личности.
Кора больших полушарий благодаря двусторонним связям с ядрами таламуса способна оказывать регулирующее воздействие на их функциональную активность.
Таким образом, основными функциями таламуса являются:
переработка сенсорной информации от рецепторов и подкорковых переключающих центров с последующей передачей её коре;
участие в регуляции движений;
обеспечение связи и интеграции различных отделов мозга .
Гипоталамус
Общее строение и расположение гипоталамуса.
Гипоталамус ( hypothalamus) представляет собой вентральный отдел (т.е. брюшной) промежуточного мозга. В его состав входит комплекс образований, расположенных под III желудочком. Гипоталамус спереди ограничивается зрительным перекрестом ( хиазмой), латерально - передней частью субталамуса, внутренней капсулой и зрительными трактами, отходящими от хиазмы. Сзади гипоталамус продолжается в покрышку среднего мозга. К гипоталамусу относят сосцевидные тела, серый бугор и зрительный перекрест. Сосцевидные тела располагаются по бокам средней линии кпереди от заднего продырявленного вещества. Это образования неправильной шаровидной формы белого цвета. Спереди от серого бугра располагается зрительный перекрёст . В нём происходит переход на противоположную сторону части волокон зрительного нерва, идущей от медиальной половины сетчатки. После перекрёста формируются зрительные тракты.
Серый бугор располагается кпереди от сосцевидных тел, между зрительными трактами. Серый бугор является полым выступом нижней стенки III желудочка, образованной тонкой пластинкой серого вещества. Верхушка серого бугра вытянута в узкую полую воронку , на конце которой находится гипофиз [ 4; 18].
Гипофиз: строение и функционирование
Гипофиз (hypophysis) - железа внутренней секреции, он располагается в специальном углублении основания черепа, "турецком седле" и при помощи ножки связан с основанием мозга. В гипофизе выделяют переднюю долю (аденогипофиз - железистый гипофиз ) и заднюю долю (нейрогипофиз ).
Задняя доля, или нейрогипофиз, состоит из нейроглиальных клеток и является продолжением воронки гипоталамуса. Более крупная доля - аденогипофиз, построена из железистых клеток. Благодаря тесному взаимодействию гипоталамуса с гипофизом в промежуточном мозге функционирует единая гипиталамо-гипофизарная система, управляющая работой всех эндокринных желез, а с их помощью - вегетативными функциями организма (рис.3).

Рисунок 3. Гипофиз и его влияние на другие эндокринные железы
В сером веществе гипоталамуса выделяют 32 пары ядер. Взаимодействие с гипофизом осуществляется посредством выделяемых ядрами гипоталамуса нейрогормонов - рилизинг-гормонов . По системе кровеносных сосудов они попадают в переднюю долю гипофиза (аденогипофиз), где способствуют высвобождению тропных гормонов, стимулирующих синтез специфических гормонов в других эндокринных железах.
В передней доле гипофиза вырабатываются тропные гормоны (тиреотропный гормон - тиреотропин, адренокортикотропный гормон - кортикотропин и гонадотропные гормоны - гонадотропины) и эффекторные гормоны (гормоны роста - соматотропин и пролактин) .
Гормоны передней доли гипофиза
Тиреотропный гормон (тиреотропин) стимулирует функцию щитовидной железы. Если удалить или разрушить гипофиз у животных, то наступает атрофия щитовидной железы, а введение тиреотропина восстанавливает ее функции.
Адренокортикотропный гормон (кортикотропин) стимулирует функцию пучковой зоны коры надпочечников, в которой образуются гормоны глюкокортикоиды. В меньшей степени выражено влияние гормона на клубочковую и сетчатую зоны. Удаление гипофиза у животных приводит к атрофии коркового слоя надпочечников. Атрофические процессы захватывают все зоны коры надпочечников, но наиболее глубокие изменения происходят в клетках сетчатой и пучковой зонах. Вненадпочечниковое действие кортикотропина выражается в стимуляции процессов липолиза, усилении пигментации, анаболическом влиянии.
Гонадотропные гормоны (гонадотропины). Фолликулостимулирующий гормон (фоллитропин) стимулирует рост везикулярного фолликула в яичнике. Влияние фоллитропина на образование женских половых гормонов (эстрогенов) небольшое. Этот гормон имеется как у женщин, так и у мужчин. У мужчин под влиянием фоллитропина происходит образование половых клеток (сперматозоидов). Лютеинизирующий гормон (лютропин) необходим для роста везикулярного фолликула яичника на стадиях, предшествующих овуляции, и для самой овуляции (разрыва оболочки созревшего фолликула и выхода из него яйцеклетки), образования желтого тела на месте лопнувшего фолликула. Лютропин стимулирует образование женских половых гормонов - эстрогенов. Однако для того чтобы этот гормон осуществил свое действие на яичник, необходимо предварительное длительное действие фоллитропина. Лютропин стимулирует выработку прогестерона желтым телом. Лютропин имеется как у женщин, так и у мужчин. У мужчин он способствует образованию мужских половых гормонов - андрогенов.
Эффекторные:
Гормон роста (соматотропин) стимулирует рост организма путем усиления образования белка. Под влиянием роста эпифизарных хрящей в длинных костях верхних и нижних конечностей происходит рост костей в длину. Гормон роста усиливает секрецию инсулина посредством соматомединов, образующихся в печени.
Пролактин стимулирует образование молока в альвеолах молочных желез. Свое действие на молочные железы пролактин оказывает после предварительного действия на них женских половых гормонов прогестерона и эстрогенов. Акт сосания стимулирует образование и выделение пролактина. Пролактин обладает также и лютеотропным действием (способствует продолжительному функционированию желтого тела и образованию им гормона прогестерона) .
Процессы в задней доле гипофиза
В задней доле гипофиза гормоны не вырабатываются. Сюда поступают неактивные гормоны, которые синтезируются в паравентрикулярном и супраоптическом ядрах гипоталамуса.
В нейронах паравентрикулярного ядра образуется преимущественно гормон окситоцин, а в нейронах супраоптического ядра - вазопрессин (антидиуретический гормон). Эти гормоны накапливаются в клетках задней доли гипофиза, где они превращаются в активные гормоны.
Вазопрессин (антидиуретический гормон) играет важную роль в процессах мочеобразования и в меньшей степени в регуляции тонуса кровеносных сосудов. Вазопрессин, или антидиуретический гормон - АДГ (диурез - выделение мочи) - стимулирует обратное всасывание (резорбцию) воды в почечных канальцах.
Окситоцин (оцитонин) усиливает сокращение матки. Ее сокращение резко усиливается, если она предварительно находилась под действием женских половых гормонов эстрогенов. Во время беременности окситоцин не влияет на матку, так как под влиянием гормона желтого тела прогестерона она становится нечувствительной к окситоцину. Механическое раздражение шейки матки вызывает отделение окситоцина рефлекторно. Окситоцин обладает способностью стимулировать также выделение молока. Акт сосания рефлекторно способствует выделению окситоцина из нейрогипофиза и выделению молока. В состоянии напряжения организма гипофиз выделяет дополнительное количество АКТГ, стимулирующего выброс адаптивных гормонов корой надпочечников .
Функциональное значение ядер гипоталамуса
В передне-боковой части гипоталамусаразличают переднюю и среднюю группы гипоталамических ядер (рис.4).

Рисунок 4. Топография ядер гипоталамуса
К передней группе относятся супрахиазматические ядра, преоптическое ядро, и самые крупные - супраоптическое и паравентрикулярное ядра.
В ядрах передней группы локализуются:
центр парасимпатического отдела (ПСНС) вегетативной нервной системы.
Стимуляция переднего отдела гипоталамуса приводит к реакциям парасимпатического типа: сужению зрачка, снижению частоты сокращений сердца, расширению просвета сосудов, падению артериального давления, усилению перистальтики (т.е. волнообразного сокращения стенок полых трубчатых органов, способствующего продвижению их содержимого к выходным отверстиям кишечника);
центр теплоотдачи. Разрушение переднего отдела сопровождается необратимым повышением температуры тела;
центр жажды;
нейросекреторные клетки, продуцирующие вазопрессин (супраоптическое ядро ) и окситоцин (паравентрикулярное ядро ). В нейронах паравентрикулярного и супраоптических ядер образуется нейросекрет, который по их аксонам перемещается в задний отдел гипофиза (нейрогипофиз), где высвобождается в виде нейрогормонов - вазопрессина и окситоцина , поступающих в кровь.
Повреждение передних ядер гипоталамуса приводит к прекращению выделения вазопрессина, вследствие чего развивается несахарный диабет . Окситоцин оказывает стимулирующее действие на гладкую мускулатуру внутренних органов, например матки. В целом от этих гормонов зависит водносолевой баланс организма.
В преоптическом ядре образуется один из рилизинг-гормонов - люлиберин, стимулирующий выработку в аденогипофизе лютеинизирующего гормона, контролирующего активность половых желез.
Супрахиазматические ядра принимают активное участие в регуляции циклических изменений активности организма - циркадианных, или суточных, биоритмов (например, в чередовании сна и бодрствования).
К средней группе гипоталамических ядер относят дорсомедиальное и вент-ромедиальное ядра, ядро серого бугра и ядро воронки.
В ядрах средней группы локализуются:
центр голода и насыщения. Разрушение вентромедиального ядра гипоталамуса приводит к избыточному потреблению пищи (гиперфагии) и ожирению, а повреждение ядра серою бугра - к снижению аппетита и резкому исхуданию (кахексии);
центр полового поведения;
центр агрессии;
центр удовольствия, играющий важную роль в процессах формирования мотиваций и психоэмоциональных форм поведения;
нейросекреторные клетки, продуцирующие рилизинг-гормоны (либерины и статины), регулирующие продукцию гипофизарных гормонов: соматостатин, соматолиберин, люлиберин, фоллиберин, пролактолиберин, тиреолиберин и др. Через гипоталамо-гипофизарную систему они оказывают влияние на ростовые процессы, скорость физического развития и полового созревания, формирование вторичных половых признаков, функции половой системы, а также на обмен веществ.
Средняя группа ядер контролирует водный, жировой и углеводный обмен, влияет на уровень сахара в крови, ионный баланс организма, проницаемость сосудов и клеточных мембран.
Задняя часть гипоталамусарасположена между серым бугром и задним продырявленным веществом и состоит из правого и левого сосцевидных тел.
В задней части гипоталамуса наиболее крупными ядрами являются: медиальное и латеральное ядра, заднее гипоталамическое ядро .
В ядрах задней группы локализуются:
центр, координирующий активность симпатического отдела (СНС) вегетативной нервной системы (заднее гипоталамическое ядро ). Стимуляция этого ядра приводит к реакциям симпатического типа: расширению зрачка, повышению частоты сокращений сердца и артериального давления, учащению дыхания и уменьшению тонических сокращений кишечника;
центр теплопродукции (заднее гипоталамическое ядро ). Разрушение заднего отдела гипоталамуса вызывает вялость, сонливость и снижение температуры тела;
подкорковые центры обонятельного анализатора. Медиальное и латеральное ядра в каждом сосцевидном теле являются подкорковыми центрами обонятельного анализатора, а также входят в лимбическую систему;
нейросекреторные клетки, продуцирующие рилизинг-гормоны, регулирующие продукцию гипофизарных гормонов .
Особенности кровоснабжения гипоталамуса
Ядра гипоталамуса получают обильное кровоснабжение. Капиллярная сеть гипоталамуса по своей разветвлённости в несколько раз больше, чем в других отделах ЦНС. Одной из особенностей капилляров гипоталамуса является их высокая проницаемость, обусловленная истонченностью стенок капилляров и их фенестрированностью ("окончатостью" - наличие промежутков - "окон" - между смежными эндотелиальными клетками капилляров (от лат. "fenestra " - окно). В результате этого в гипоталамусе слабо выражен гематоэнцефалический барьер (ГЭБ), и нейроны гипоталамуса способны воспринимать изменения состава спинномозговой жидкости и крови (температуру, содержание ионов, наличие и количество гормонов и т.д.).
Функциональное значение гипоталамуса
Гипоталамус является центральным звеном, связующим нервные и гуморальные механизмы регуляции вегетативных функций организма. Управляющая функция гипоталамуса обусловлена способностью его клеток к секреции и аксональному транспорту регуляторных веществ, которые переносится в другие структуры мозга, спинномозговую жидкость, кровь или в гипофиз, изменяя функциональную активность органов-мишеней.
В гипоталамусе выделяют 4 нейроэндокринные системы:
Гипоталамо-экстрагипоталамная система представлена нейросекреторными клетками гипоталамуса, аксоны которых уходят в таламус, структуры лимбической системы, продолговатый мозг. Эти клетки выделяют эндогенные опиоиды, соматостатин и др.
Гипоталамо-аденогипофизарная система связывает ядра заднего гипоталамуса с передней долей гипофиза. По этому пути транспортируются рилизинг-гормоны (либерины и статины). Посредством их гипоталамус регулирует секрецию тропных гормонов аденогипофиза, определяющих секреторную активность желёз внутренней секреции (щитовидной, половых и др.).
Гипоталамо-метагипофизарная система связывает нейросекреторные клетки гипоталамуса с гипофизом. По аксонам этих клеток транспортируются меланостатин и меланолиберин, которые регулируют синтез меланина - пигмента, определяющего окраску кожи, волос, радужки и других тканей организма.
Гипоталамо-нейрогипофизарная система связывает ядра переднего гипоталамуса с задней (железистой) долей гипофиза. По этим аксонам транспортируются вазопрессин и окситоцин, которые накапливаются в задней доле гипофиза и выделяются в кровоток по мере необходимости .
Заключение
Таким образом, дорсальный отдел промежуточного мозга представляет собой филогенетически более молодой таламический мозг, являющийся высшим подкорковым сенсорным центром, в котором переключаются практически все афферентные пути, несущие сенсорную информацию от органов тела и органов чувств к большим полушариям головного мозга. К задачам гипоталамуса относится также управление психоэмоциональным поведением и участие в реализации высших психических и психологических процессов, в частности памяти.
Вентральный отдел - гипоталамус являетсяболее старым в филогенетическом отношении образованием. Гипоталамо-гипофизарная система осуществляет контроль над гуморальной регуляцией водносолевого баланса, обменом веществ и энергии, работой иммунной системы, терморегуляцией, репродуктивной функцией и т.д. Выполняя и этой системе регулирующую роль, гипоталамус является высшим центром, управляющим автономной (вегетативной) нервной системой.
Список литературы
1. Анатомия человека / Под ред. М.Р. Сапина. - М.: Медицина, 1993.
2. Блум Ф., Лейзерсон А., Хофстедтер Л. Мозг, разум поведение. - М.: Мир, 1988.
3. Гистология / Под ред. В.Г. Елисеева. - М.: Медицина, 1983.
4. Привес М.Г., Лысенков Н.К., Бушкович В.И. Анатомия человека. - М.: Медицина, 1985.
5. Синельников Р.Д., Синельников Я.Р. Атлас анатомии человека. - М.: Медицина, 1994.
6. Тишевской И.А. Анатомия центральной нервной системы: Учебное пособие. - Челябинск: Изд-во ЮУрГУ, 2000.







