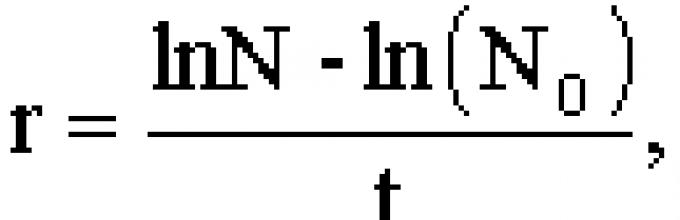
Рост популяции
Рост популяции
- изменение численности ее особей от момента размножения и вселения особей в биоценоз. Определяется как прирост численности в единицу времени (см. индекс роста популяции). Рост популяции может быть выражен J-образной (экспоненциальной) или S-образной (логистической) кривыми роста. В первом случае кривую отражает уравнение dN/dt=rN (с определенным пределом для N), во втором -dN/dt=rN(k-N)/k, где N - численность популяции; dN и dt - изменение численности и времени; r - потенциальный рост, а k - предел роста (рис. 28). Практически кривые имеют массу вариантов, принимающих характер флюктуации.
Экологический словарь. - Алма-Ата: «Наука» . Б.А. Быков . 1983 .
- Рождаемость
- Рудеральные организмы
Смотреть что такое "Рост популяции" в других словарях:
Или длина тела человека расстояние от верхушечной точки головы до плоскости стоп. В антропологии рост является одним из общих антропометрических признаков. Входит в список показателей физического развития человека. На рост человека, в числе … Википедия
РОСТ ЗАТУХАЮЩИЙ - замедляющийся (до полного прекращения) рост популяции (при достижении предела асимптоты верхней), после чего устанавливается колебательное равновесие. Затухающий рост описывается логистической кривой, имеющей S образную форму. Замедление… … Экологический словарь
Увеличение массы и линейных размеров индивидуума (особи) и его отд. органов, происходящее за счёт увеличения числа и массы клеток, а также неклеточных образований в результате преобладания процессов анаболизма над процессами катаболизма. У… … Биологический энциклопедический словарь
Рост. В микробиологии понятие Р. используют в 2 случаях: 1) Р. клеток процесс увеличения биомассы (размеров, объема, массы) организма или его частей в ходе индивидуального развития, обусловленный синтезом новых веществ. Характерен для всех групп… … Словарь микробиологии
РОСТ - означает увеличение размеров развивающегося организма. В типичных случаях Р. связан с увеличением массы, однако не всякое увеличение массы организма мы, обозначаем как Р. (напр. отложение жира, накопление половых продуктов у некоторых животных,… … Большая медицинская энциклопедия
Увеличение массы индивидуума (особи), происходящее за счёт увеличения числа клеток, массы клеток и неклеточных образований. Р. живой системы происходит в результате преобладания Анаболизма над Катаболизмом. У животных в процессе… … Большая советская энциклопедия
СТАЦИОНАРНОЕ СОСТОЯНИЕ ПОПУЛЯЦИИ - фаза роста популяции, при которой рождаемость равняется смертности и плотность популяции сохраняется на постоянном уровне. Стационарное состояние популяции достигается при переходе отрицательного ускорения роста популяции в верхнюю асимптоту,… … Экологический словарь
ПОКАЗАТЕЛЬ ПОТЕНЦИАЛЬНОГО РОСТА ПОПУЛЯЦИИ - специфическая скорость роста популяции в условиях стационарного и стабильного распределения возрастов rmax. Часто максимальную величину r называют биотическим, или репродуктивным, потенциалом. См. также Рост популяции. Экологический… … Экологический словарь
Плотность популяции, число особей (животных, растений, микроорганизмов) в расчёте на единицу объёма (воды, воздуха или почвы) или поверхности (почвы или дна водоёма). Плотность популяции важный экологический показатель пространственного… … Википедия
Плотность популяции число особей (животных, растений, микроорганизмов) в расчете на единицу объема (воды, воздуха или почвы) или поверхности (почвы или дна водоема). Плотность популяции важный экологический показатель… … Википедия
Книги
- Активное долголетие , Герман Клименко. «Активное долголетие» – тематический альманах Радио «Медиаметрикс». Увеличение продолжительности жизни, рост численности пожилых людей в общей популяции населения, рост доли нетрудоспособного…
Любая популяция теоретически способна к неограниченному росту численности, если ее не лимитируют факторы внешней среды. Даже самый медленно размножающийся вид способен в короткое время произвести столько особей, что для них не хватит места на земном шаре. Всего за пять поколений, т. е. за 1–1,5 летних месяца, одна единственная тля может оставить более 300 млн. потомков. Если бы все зародыши сохранялись, а все потомство выживало, численность любой популяции через определенные интервалы увеличивалась бы в геометрической прогрессии , и это несмотря на то, что одни производят за всю жизнь всего несколько яиц или детенышей, а другие – тысячи и даже миллионы зародышей. Рост в геометрической прогрессии выражается на графике экспоненциальной кривой . В.И. Вернадский назвал этот процесс «давлением жизни» .
Фактически во всех живых организмах заложена способность размножаться беспредельно. Эта способность делает жизнь очень мощной силой на Земле. Огромная живая масса организмов поддерживает круговорот веществ на планете, создает горные породы, почвы, регулирует состав вод и атмосферы. Но в природе биотический потенциал популяции никогда не реализуется полностью. Главный ограничитель на пути к беспредельному размножению организмов – нехватка ресурсов , самых необходимых: для растений – минеральных солей, углекислого газа, воды, света; для животных – пищи, воды; для микроорганизмов – разнообразных потребляемых ими соединений. Запасы этих ресурсов не бесконечны, в разных частях планеты они имеют свои пределы, и этим сдерживается размножение видов. Второй ограничитель – влияние неблагоприятных условий , замедляющих рост и размножение организмов, даже если есть необходимые для этого ресурсы. Наконец, гибель зародышей или подрастающих молодых особей от врагов и болезней . Так, тысячи желудей, которые ежегодно производит один большой дуб, оказываются съеденными белками, кабанами, сойками, мышами, насекомыми, или поражаются плесневыми грибками и бактериями, или гибнут на стадии проростков по разным причинам. В результате лишь из считанных желудей вырастают взрослые деревья.
Общие изменения численности популяции складываются за счет четырех явлений: рождаемости, смертности, вселения и выселения особей (иммиграция и эмиграция).
Рождаемость (скорость рождаемости) – число новых особей, появившихся в популяции за единицу времени в результате размножения.
Различают максимальную и фактическую рождаемость. Максимальная рождаемость – максимальная реализация возможности рождения при отсутствии лимитирующих факторов среды. Фактическая рождаемость – реальная реализация возможности рождения.
Различают абсолютную и удельную рождаемость. Абсолютная (общая) рождаемость, или скорость рождаемости – число особей (яиц, семян и т. п.), родившихся (отложенных, продуцированных и т. д.) за некоторый промежуток времени.
Удельная рождаемость – отношение скорости рождаемости к исходной численности. Эта величина зависит от интенсивности размножения особей: для бактерий – час, для фитопланктона – сутки, для насекомых – неделя или месяц, для крупных млекопитающих – год.
Величина рождаемости зависит от многих причин. Большое значение имеет доля особей, способных в данный период к размножению, что определяется соотношением полов и возрастных групп. Важна также частота последовательности генераций. Так, среди насекомых различают моновольтинные и поливольтинные виды. Первые дают одну, вторые – несколько генераций за год. Например, у тлей за сезон насчитывается до 15 партеногенетических поколений. По числу периодов размножения в течение жизни различают моноциклические и полициклические виды. Моноцикличность, или однократное размножение, свойственна обычно видам с короткой продолжительностью жизни в половозрелом состоянии (лососевые рыбы, поденки, майские жуки и многие другие насекомые). Полицикличность характеризуется повторным размножением особей и присуща большинству позвоночных животных и ряду беспозвоночных, например ракообразным.
У растений выделяют монокарпические и поликарпические виды, т. е. с однократным и многократным размножением в течение жизни.
Размножаемость популяции, как правило, не бывает прямо пропорциональна плодовитости. Плодовитость сильно зависит от степени развития заботы о потомстве или обеспеченности яиц питательными материалами. Среди рыб наибольшее количество икринок выметывают виды с пелагической икрой – сельди, тресковые, камбаловые и др. Например, сахалинская сельдь продуцирует 38–46 тыс. мелких, в доли миллиметра, икринок. У лососевых, зарывающих икру в грунт, развивается меньшее число яиц, но более крупных размеров. Средняя плодовитость амурской горбуши 1300–1500 икринок диаметром 4–6 мм. Наиболее крупная икра у акул и химер, до 6–8 см. Яйца этих рыб имеют к тому же плотную защитную оболочку. Количество их очень невелико – несколько штук на одну самку.
У видов, оберегающих и выкармливающих свой молодняк, плодовитость резко понижена. Размер кладки у птиц разных видов варьирует уже не в тысячи раз, как у рыб, а в пределах от одного яйца (у некоторых хищников, пингвинов, чистиков и др.) до 20–25 (максимальное количество яиц у куриных, например, у серой куропатки).
Большая плодовитость вырабатывается у видов также в условиях более высокой смертности, особенно при сильном давлении со стороны хищников. Отбор на плодовитость компенсирует высокую норму гибели в популяциях. Поэтому при высокой плодовитости рост численности популяции может быть весьма низким. В различных популяциях одного вида плодовитость обычно тем выше, чем более неблагоприятны занимаемые ими места обитания. Так, у многих млекопитающих – зайцев, мышей, полевок – число детенышей в помете на границах ареала больше, чем в его центре.
Гипотеза дифференцированной специализации полов (В. А. Геодокян) – дифференциация полов происходит по двум основным направлениям эволюции: изменению (мужской пол) и сохранению (женский пол). Чем больше в популяции женских особей, тем лучше сохраняется ее генотипическая структура; чем больше мужских особей, тем больше скорость или величина изменения этой структуры.
Смертность (скорость смертности) – число особей, погибших в популяции за единицу времени (от хищников, болезней, старости и других причин). Смертность – величина обратная рождаемости.
Различают минимальную и фактическую смертность. Минимальная смертность – минимально возможная величина смертности. Фактическая смертность – реальная величина смертности.
Различают абсолютную и удельную смертность. Абсолютная (общая) смертность, или скорость смертности – число особей, погибших за промежуток времени.
Удельная смертность (а) – отношение скорости смертности к исходной численности.
Скорость роста популяции – изменение численности популяции за единицу времени. Скорость роста популяции может быть положительной, нулевой и отрицательной. Она зависит от показателей рождаемости, смертности и миграции (вселения – иммиграции и выселения – эмиграции). Увеличение (прибыль) численности происходит в результате рождаемости и иммиграции особей, а уменьшение (убыль) численности – в результате смертности и эмиграции особей.
Различают абсолютную и удельную скорость роста популяции. Абсолютная (общая) скорость роста – изменение численности популяции за промежуток времени.
Удельная скорость роста – отношение скорости роста к исходной численности.
При отсутствии лимитирующих факторов среды удельная скорость роста равна величине, которая характеризует свойства самой популяции и называется удельной (врожденной) скоростью роста популяции или биотическим потенциалом вида.
Величина биотического потенциала очень различается у разных видов. Например, самка косули способна произвести за жизнь 10–15 козлят, трихина отложить 1,8 тыс. личинок, самка медоносной пчелы – 50 тыс. яиц, рыба-луна – до 3 млрд. икринок.
Однако в природе, в связи с действием лимитирующих факторов, биотический потенциал популяции никогда не реализуется полностью. Его величина обычно складывается как разность между рождаемостью и смертностью в популяции:
где b – число родившихся, d – число погибших особей в популяции за один и тот же период времени.
Когда b = d, r = 0 и популяция находится в стационарном состоянии. Когда b > d, r > 0, численность популяции увеличивается. Когда b < а, r < 0, численность популяции сокращается. Формула d = b – r позволяет определить смертность, которую трудно измерить непосредственно, а определить r достаточно просто непосредственными наблюдениями.
Скорость роста может быть выражена в виде кривой роста популяции. Существуют две основные модели роста популяции: J- образная и S -образная.
J-образная кривая (1) отражает неограниченный экспоненциальный рост численности популяции, не зависящий от плотности популяции. Такой тип роста возможен пока биотический потенциал популяции (r ) реализуется полностью. Это продолжается, пока низка конкуренция за ресурсы. Однако после превышения емкости среды (предельной плотности насыщения, предельной численности) (К ), произойдет резкое снижение численности.
S-образная (сигмоидная, логистическая) кривая (2) отражает логистический тип роста, зависящего от плотности популяции, при котором скорость роста популяции снижается по мере роста численности (плотности). Скорость роста снижается вплоть до нуля при достижении предельной численности.
Если среда не оказывает ограничивающего влияния, то специфическая скорость роста популяции для данных микроклиматических условий постоянна и максимальна. В таких благоприятных условиях скорость роста характерна для некоторой возрастной структуры популяции и служит единственным показателем наследственно обусловленной способности популяции к росту. Она является экспонентой в дифференциальном уравнении роста популяции в нелимитированной среде при специфических физических условиях:
dN / dt = rN; r = dN / (Ndt);
ln n t = rt - ln(N 0); n t = N 0 e rt ,
где N 0 - численность в нулевое время, n t - численность в момент времени t, e - основание натуральных логарифмов.
Из уравнения ln n t = t - ln(N 0) можно вычислить скорость роста популяции:
![]()
Показатель r - фактическая разность между специфической мгновенной скоростью рождений (b) и специфической мгновенной скоростью гибели организмов (d). Она может быть выражена в виде:
r = b - d.
Общая скорость роста популяции в отсутствие лимитирующего влияния среды зависит от возрастного состава и вклада в репродукцию различных возрастных групп. Следовательно, вид может характеризоваться величинами r в зависимости от структуры популяции. При определении стационарного и стабильного распределения возрастов специфическую скорость роста называют показателем потенциального роста популяции (r max). Часто этот показатель называют биотическим, или репродуктивным, потенциалом (термин введен Р. Чепменом в 1928 г.). Разность между биотическим потенциалом и фактической скоростью роста считают мерой сопротивления среды, которая характеризует сумму всех лимитирующих факторов, препятствующих реализации биотического потенциала.
Помимо приведенных, используют и другие уравнения роста популяции (Р.Уиттекер, 1980). При неограниченном росте численности популяции можно использовать геометрические и экспоненциальные формулы.
Геометрические формулы:
N 1 / N 0 = R - темп роста популяции за единицу времени;
N 1 = N 0 R t - плотность популяции через время t.
Экспоненциальные формулы:
dN / dt = Nn - скорость роста популяции;
Nt = N 0 e rt - плотность популяции через время t.Математический метод можно использовать и при описании неограниченного роста популяции. В случае ограниченного роста популяции скорость ее роста и плотность можно вычислять по следующим логистическим формулам:
![]() - скорость роста;
- скорость роста;

где N - плотность популяции; N 0 - начальная плотность популяции; N 1 - плотность популяции через единицу времени роста популяции (t= 1); N t - плотность популяции через время t при постоянной скорости роста, К - емкость среды для максимальной плотности популяции, е - основание натурального логарифма. Величина К называется предельной допустимой нагрузкой на среду, или емкостью среды для данной популяции.
Выбор математической модели определяется задачами исследований и адекватностью модели для каждого определенного случая.
Типы роста популяций. Представление о емкости местообитания. В зависимости от характера роста численности популяций выделяют различные типы их роста. По форме кривых, построенных на арифметической шкале, можно выделить два основных типа роста, описываемых J-образной и S-образной кривыми. Эти два типа кривых могут модифицироваться различным образом (рис. 18).

Рис. 18. Кривые роста численности популяции (по Ю. Одуму, 1875)
А - J-образная, В - S-образная форма кривой роста численности популяции. A 0 - вначале наблюдается неограниченный рост численности популяции; а 1 - вначале наблюдается неограниченный рост численности, затем он прекращается и при благоприятных условиях вновь возобновляется, достигая прежней величины; А 2 - наблюдается нелимитированный рост численности, затем он внезапно прекращается и дальше наблюдаются колебания на более низком уровне; В 0 - происходит рост численности популяции по S-образной кривой, достигая К-уровня; В 2 - вначале наблюдается медленный рост численности, затем скорость возрастает и достигает К-уровня; В 1 - при достижении К-уровня наблюдаются небольшие отклонена от него; В 3 - наблюдаются значительные отклонения от К-уровня.
При J-образной форме кривой роста плотность быстро увеличивается, но затем, когда начинает действовать лимитирующий фактор, рост популяции внезапно прекращается. Такой тип роста может быть описан экспоненциальным уравнением:
Уравнение J-образной кривой то же, что и при определении скорости роста. Разница состоит в том, что величина N имеет предел. Это означает, что относительно неограниченный рост внезапно останавливается, когда популяция исчерпает свои pecypcы (пища, жизненное пространство) или вмешается какой-либо друге фактор. После того как верхний предел N будет достигнут, плотность некоторое время может оставаться на этом уровне либо резко падать. Это характерно для природных популяций насекомых, водорослей и др.
При логистической (S-образной) кривой роста популяции вначале увеличение численности идет очень медленно, затем быстрее, но потом под влиянием факторов сопротивления среды рост популяции постепенно замедляется. Это замедление, обусловливаемо сопротивлением среды, становится все более выраженным и в конечном итоге достигает определенной величины. Затем начинае поддерживаться более или менее устойчивое равновесие. Такой тип роста может быть выражен уравнением Ферхюльста-Пирла:
![]()
где К - константа, обозначающая верхний предел увеличения численности популяции, называемая верхней асимптотой для S-образно кривой.
Характерная форма S-образной кривой обусловлена постепенным усилением действия неблагоприятных факторов по мере увеличения плотности популяции. Такой тип роста отличается от J-образного, при котором популяция начинает испытывать давление среды в конце роста.
В простом, или идеальном, случае усиление действия неблагоприятных факторов в зависимости от плотности популяции является линейным и его можно записать следующим образом:
![]()
где r - скорость роста популяции или показатель потенциального роста, N- величина популяции, К- максимально возможная величина популяции; е - основание натуральных логарифмов, а - константа интегрирования, определяющая положение кривой относительно начала координат, t - время.
Это уравнение отличается от экспоненциального тем, что содержит выражения (К - N) / К, (r / K) N 2 или (1 - N) / К. Эти выражения представляют три показателя, характеризующих сопротивление среды, создаваемое вследствие роста популяции, которая по мере приближения к пределу уменьшает скорость потенциальной репродукции. Это уравнение отражает закон: скорость увеличения популяции равна максимально возможной скорости роста популяции, умноженной на степень реализации максимальной скорости.
Следует отметить, что для описания изменений численности популяции существует много математических уравнений, решение которых можно представить графически в виде S-образных кривых. Это справедливо почти для любого уравнения, в котором увеличение отрицательных факторов находится в какой-либо зависимости от плотности популяции.
Для сравнения экспериментальных данных с теоретической кривой следует убедиться в том, что показатели, входящие в уравнение, характеризуют воздействия, регулирующие плотность популяции. Ситуации, когда сопротивление среды возрастает линейно при увеличении плотности, могут иметь место в популяциях с простым жизненным циклом. В более высокоорганизованных популяциях, со сложным биологическим циклом и длительными периодами индивидуального развития, изменения, вероятно, отсрочены во времени.
Стратегии развития популяций. В зависимости от типа кривой роста численности популяции выделяют стратегию развития, определяемую такими свойствами, как скорость размножения, характер передачи энергии от одного поколения другому, колебания численности относительно равновесного значения, или К-уровня, скорость изменения численности, приспособленность вида к конкретной территории, размеры особей, продолжительность их жизни и т.д. (табл. 9).
Таблица 9. Характерные особенности r- и K-видов.
| r-виды | К-виды |
| Размножаются быстро (высокая плодовитость, короткое время генерации), поэтому значение r (врожденная скорость роста популяции) высокое | Размножаются медленно (низкая плодовитость, продолжительное время генерации), поэтому значение r низкое |
| Скорость размножения не зависит от плотности популяции | Скорость размножения зависит от плотности популяции быстро увеличивается, если плотность падает |
| Энергия и вещество распределяются между многими потомками | Энергия и вещество концентрируются у немногих потомков, родители заботятся о потомстве |
| Размеры популяции некоторое время могут превышать К-уровень | Размеры популяции близки к равновесному значению, определяемому К-уровнем |
| Вид не всегда устойчив на данной территории | Вид устойчив на данной территории |
| Расселяются широко и в больших количествах, у животных может мигрировать каждое поколение | Расселяются медленно |
| Размножение идет с относительно большими затратами энергии и вещества | Размножение идет с относительно мапыми затратами энергии и вещества большая часть энергии и вещества расходуется на репродуктивный (вегетативный) рост |
| Малые размеры особей | Крупные размеры особей, у растений деревянистые стебли и большие корни |
| Малая продолжительность жизни особи | Большая продолжительность жизни особи |
| Могут поселяться на открытом грунте | Плохо приспособлены к росту на открытых местах |
| Местообитания сохраняются недолго | Местообитания устойчивы и сохраняются долго |
| Слабые конкуренты | Сильные конкуренты |
| Защитные приспособления развиты сравнительно слабо | Хорошие защитные механизмы |
| Не становятся доминантами | Могут становиться доминантами |
| Лучше приспособлены к изменениям окружающей среды (менее специализированные) | Менее устойчивы к изменениям ycловий среды (высокая специализация для жизни в устойчивых местообитаниях) |
Для характеристики стратегии популяции используют символы r и К. Быстро размножающиеся виды с высоким значением r называются K-видами. К ним относятся типичные пионерные виды нарушенных местообитаний.
Виды с относительно низким значением г называются к-видами. Скорость их размножения в значительной степени зависит от плотности популяции и близка к равновесному значению, определяемому -уровнем. Эти виды характерны для поздних стадий развития сукцессии.
Следует отметить, что существует ряд промежуточных стратегий. Эти две стратегии популяций представляют два способа решения одной задачи - задачи выживания вида. Виды, относящиеся к г-стратегии, быстрее, чем виды, относящиеся к К-стратегии, заселяют нарушенные местообитания, характерные для ранних сукцессий (обнаженнaя горная порода, лесные вырубки, бывшие карьеры), так как они легче распространяются и быстрее размножаются. Виды с К-стратегией более конкурентоспособны и в конечном итоге вытесняют виды с r-стратегией, которые могут перемещаться в другие нарушенные местообитания. Так как виды с r-стратегией обладают высоким репродукционным потенциалом, то это означает, что, оставшись в каком-либо местообитании, они быстро использовали бы доступные ресурсы и превысили бы поддерживающую емкость среды, что привело бы к гибели популяции. Для видов с r-стратегией характерна J-образная кривая роста с быстрым падением численности популяции.
СКОРОСТЬ РОСТА ПОПУЛЯЦИИ
прирост популяции за единицу времени: D
N/D
t, где N - размер популяции (численность), t - время. Может быть постоянной (неограниченный рост - при условии отсутствия сопротивления среды приросту) или затухающей (когда с увеличением численности особей условия их существования ухудшаются и темп воспроизводства падает).
- - allopatric populations - .Популяции одного вида, обитающие в разных географических районах и не соприкасающиеся друг с другом...
Молекулярная биология и генетика. Толковый словарь
- - прирост массы растения или его отдельного органа в единицу времени...
Словарь ботанических терминов
- - существование двух обл. равновесия в популяции при неизменных условиях среды, возникающих в результате сильной зависимости свойств самой популяции от ее численности...
Экологический словарь
- - численность или плотность популяции на определенной территории, в определенной экологической...
Экологический словарь
- - специфическая скорость роста популяции в условиях стационарного и стабильного распределения возрастов - rmax. Часто максимальную величину r называют биотическим, или репродуктивным,...
Экологический словарь
- - скорость роста животных, определяемая по формуле И. И. Шмальгаузена и С. Броди: где ln - масса животного в конце опыта; lo - масса животного в начале опыта; tn-to - время опыта...
Экологический словарь
- - определяется по уравнению: где N - численность популяции; t - время; r - максимальная специфическая скорость увеличения популяции; K - емкость среды, или предельная плотность насыщения...
Экологический словарь
- - степени продолжительности перевозки грузов по железным дорогам...
Справочный коммерческий словарь
- - скорость изменения числа организмов r в зависимости от времени в данный момент. Определяется по формуле: ║ где dN - изменение числа организмов в данный момент времени dt...
Экологический словарь
- - мера мгновенной удельной скорости изменения размера популяции; выражается числом особей на единицу времени и на одну особь, т. е. имеет размерность 1/время...
Экологический словарь
- - способность популяции предотвращать потери, возникающие в случае, когда среда становится неблагоприятной. Термин предложил Р. Уиттекер...
Экологический словарь
- - скорость увеличения численности популяции за год. Определяется по формуле: , где R - чистая скорость размножения; Т - среднее время генерации...
Экологический словарь
- - : Ir=log R0/Tc, где R0 - скорость воспроизводства, во сколько раз увеличивается численность популяции за одну генерацию; Тс - средний возраст первого размножения или плодоношения...
Экологический словарь
- - параметр λ, скорость увеличения численности популяции в расчете на 1 особь и на единицу времени...
Экологический словарь
- - графическое изображение роста популяции. См. Логистическая кривая...
Экологический словарь
- - см. Скорость размножения чистая...
Экологический словарь
"СКОРОСТЬ РОСТА ПОПУЛЯЦИИ" в книгах
14 Популяции
автора Барнетт ЭнтониРастущие популяции
Из книги Род человеческий автора Барнетт ЭнтониРастущие популяции Если мы рассмотрим теперь влияние рождаемости и смертности на численность населения, то обнаружим любопытную тенденцию: население земного шара постоянно увеличивается. При сохранении современных темпов роста к 1980 г. население Земли достигнет 4
Уменьшающиеся популяции?
Из книги Род человеческий автора Барнетт ЭнтониУменьшающиеся популяции? Несмотря на низкий процент смертности, популяции Западной Европы и британских доминионов почти не изменяются: понизившийся уровень рождаемости уравновешивает снижение смертности. Если бы не такое «равновесие», уровень жизни в этих странах не
Популяции будущего
Из книги Род человеческий автора Барнетт ЭнтониПопуляции будущего Каждую популяцию можно рассматривать как состояние в ряду изменений, зависящих от соотношения между собою рождаемости и смертности. Поэтому демографическая политика страны должна строиться с учетом состояния в данное время возможных изменений в
Глава 8. ПОПУЛЯЦИИ
Из книги Общая экология автора Чернова Нина МихайловнаГлава 8. ПОПУЛЯЦИИ 8.1. Понятие о популяции в экологии Популяцией в экологии называют группу особей одного вида, находящихся во взаимодействии между собой и совместно населяющих общую территорию.Слово «популяция» происходит от латинского «популюс» – народ, население.
8.5.6. Темпы роста популяции
Из книги Общая экология автора Чернова Нина МихайловнаРОСТ ПОПУЛЯЦИИ
Из книги Экология автора Митчелл ПолРОСТ ПОПУЛЯЦИИ В каком-то из изданий было сказано, что если бы человеческая популяция продолжала расти с нынешней скоростью, то через 200 лет огромная масса людей устремилась бы в космос со скоростью света. Этого, конечно, не произойдет; это всего лишь шутка, показывающая,
14 Популяции
Из книги Род человеческий автора Барнетт Энтони14 Популяции Плодитесь и размножайтесь, и наполняйте землю, и обладайте ею, и владычествуйте над рыбами морскими, и над птицами небесными, и над всяким животным, пресмыкающимся по земле. Бытие Хотя голод и болезни - эти извечные враги человечества - косили людей с
8.5.6. Темпы роста популяции
Из книги Общая экология автора Чернова Нина Михайловна8.5.6. Темпы роста популяции Графики роста числа членов любой природной популяции в новых для нее условиях сильно отличаются от экспоненты. Кривая после подъема разной степени крутизны поворачивает параллельно горизонтальной оси, чем знаменует установление некоторой
Глава 27 Большие ожидания (роста) Пределы корпоративного роста
Из книги Больше, чем вы знаете. Необычный взгляд на мир финансов автора Мобуссин МайклГлава 27 Большие ожидания (роста) Пределы корпоративного роста В воздушных замках легко найти убежище. И строить их так легко. Генрик Ибсен. Строитель Сольнес Я слышу все больше прогнозов насчет высоких темпов роста прибыли в будущем. Кое-кто более сдержан в своих
1.6. Может ли скорость обмена информацией превышать скорость света?
Из книги Квантовая магия автора Доронин Сергей Иванович1.6. Может ли скорость обмена информацией превышать скорость света? Довольно часто приходится слышать, что эксперименты по проверке неравенств Белла, опровергающие локальный реализм, подтверждают наличие сверхсветовых сигналов. Это говорит о том, что информация способна
Скорость роста толстолобика
Из книги Ловля карпа, леща, толстолобика, белого амура. Секреты и хитрости успешной рыбалки автора Сторожев КонстантинСкорость роста толстолобика При благоприятном температурном режиме и хорошей кормовой базе толстолобики растут очень быстро. Сеголетка может весить до 1 кг. В возрасте 2 лет пестрый толстолобик достигает массы 2–2,5 кг, белый – 1,5–2 кг. Взрослый же толстолобик (5–6 лет и
Скорость тренировочного чтения должна в три раза превышать скорость обычного чтения
Из книги Скорочтение. Как запоминать больше, читая в 8 раз быстрее автора Камп ПитерСкорость тренировочного чтения должна в три раза превышать скорость обычного чтения Основное правило тренировок заключается в том, что если вы хотите читать с определенной скоростью, то вам нужно выполнять тренировочное чтение приблизительно в три раза быстрее. Так,
Параграф 29. Стимуляция роста юных спортсменов «программа роста»
Из книги С самого начала (путь тренера) автора Головихин Евгений ВасильевичПараграф 29. Стимуляция роста юных спортсменов «программа роста» В самом начале тренерского пути я обратил внимание на рослых и мощных спортсменов, которые обладали анатомическими способностями с рождения. Эти спортсмены сразу имели преимущество перед сверстниками. Они
51. Скорость истечения в сужающемся канале, массовая скорость перемещения потока
Из книги Теплотехника автора Бурханова Наталья51. Скорость истечения в сужающемся канале, массовая скорость перемещения потока Скорость истечения в сужающемся каналеРассмотрим процесс адиабатного истечения вещества. Предположим, что рабочее тело с некоторым удельным объемом (v1) находится в резервуаре под
Чтобы получить полную картину динамики численности той или иной популяции, а также рассчитать скорость ее роста, наряду с данными о том, как распределяется по разным возрастам смертность, необходимо знать также, в каком возрасте особи начинают производить потомство и какова средняя плодовитость особей разного возраста. Поэтому в таблицы для расчета скорости роста популяции к графам, характеризующим выживаемость, добавляют графу, в которой записывают среднее число потомков, появившихся в течение данного возрастного интервала в расчете на одну особь родительского поколения. Для простоты представим себе гипотетический пример животного, начинающего размножаться на третьем году жизни и живущего, как правило, не более 10 лет. В первой графе табл. 4 запишем возраст (х), во второй - долю особей, доживших до данного возраста от начальной численности когорты (l х), в третьей-среднее число потомков, появившихся на свет у особей данного возраста в расчете на одну родительскую особь (m х), в четвертой-произведение доли доживших особей на среднюю их плодовитость (l x m x). Сумма последних величин по всему столбцу есть величина, называемая чистой скоростью воспроизводства. Чистая скорость воспроизводства показывает, во сколько раз увеличивается численность популяции за одно поколение. Если R 0 = 1, то популяция стационарна-численность ее сохраняется постоянной, поскольку каждое последующее поколение точно замещает предыдущее. В демографии обычно составляют отдельные таблицы для женщин (тогда в графе m х - среднее число дочерей, родившихся от матерей денного возраста) и для мужчин (в графе т х - среднее число сыновей, появившихся у отцов данного возраста).
Величина R 0 сильно варьируется в зависимости от вида организмов, а также условий его существования. Так, например, для содержавшейся в хороших лабораторных условиях популяции пашенной полевки Microtus agrestis величина R 0 оказалась равной 5,90, а у лабораторной популяции рисового долгоносика Саlandra oryzae - 113,48. Таким образом, за одно поколение в благоприятных условиях популяция пашенной полевки может увеличить свою численность примерно в 6 раз, а популяция рисового долгоносика - в 113 раз.
| x | l x | m x | l x m x |
| 1,00 | |||
| 0,60 | |||
| 0,50 | |||
| 0,45 | 2,0 | 0,90 | |
| 0,40 | 2,5 | 1,0 | |
| 0,37 | 1,5 | 0,55 | |
| 0,33 | 1,0 | 0,33 | |
| 0,20 | 0,5 | 0,10 | |
| 0,10 | 0,1 | 0,01 | |
| 0,05 | |||
| 0,00 | |||
| Σl x m x = 2.89 |
Уже из самого способа расчета R 0 ясно, что величина эта определяется комбинацией выживаемости (l x -кривой) и плодовитости (m x -кривой). Перемножая значения l х и m x для каждого возраста х, мы тем самым определяем площадь под кривой l х m х. Так же как и l х, величина m x может сильно меняться в зависимости от вида организмов и условий его существования.
Использовать показатель R 0 при сравнении видов, характеризующихся разной продолжительностью жизни, не всегда удобно. Гораздо лучше употреблять в таких случаях величину r показатель специфической скорости роста популяции. Чтобы установить связь между этими величинами, представим себе, что в течение промежутка времени, равного длительности одного поколения (= времени генерации) Т, популяция растет экспоненциально. Тогда численность популяции к концу временного интервала Т будет равной N T = N 0 e rT .. Из последнего уравнения следует, что N T /N 0 = e rT . Но ведь N T /N 0 есть не что иное, как отношение численности особей в двух следующих друг за другом поколениях, или, другими словами, величина R 0 - Переписав это уравнение в несколько иной форме: R 0 = e rT , мы можем определить из него и величину r по формуле r = lnR 0 /T .
Приведенный способ оценки показателя r точен настолько, насколько точно определена длительность поколения T . В некоторых случаях вопрос о том, что такое длительность поколения, решается достаточно просто. Так, для некоторых лососевых рыб, например горбуши (Oncorhynchus gorbuscha) или нерки (Опсоrhynchus nerka ), мечущих икру один раз в конце жизни и после того погибающих, длительность поколения - это, очевидно, время от откладки икры (или выклева из икры личинок) до размножения выросших из этих икринок (личинок) особей. Подобным образом раз в конце жизни происходит размножение у многих насекомых (достаточно вспомнить поденок) и ряда видов растений. Однако у многих животных и растений период размножения растянут во времени, причем в пределах того возраста, когда размножение возможно, среднее число потомков на родительскую особь меняется. В этом случае величину длительности поколения приближенно можно рассчитать следующим образом:
Смысл подобного способа расчета легко уяснить, обратившись к механической модели (Dublin, Lotka, 1925), иллюстрирующей реальный пример из человеческой популяции. Представим себе шкалу возраста матери (рис. 25) в виде горизонтальной планки, установленной как балансир на одной опоре в центре (по типу качелей из доски, положенной на бревно). Начало отсчета (момент рождения матери) соответствует точке опоры, от которой идут симметричные шкалы влево и вправо по плечам балансира. На правое плечо нанесена гистограмма, показывающая число дочерей, родившихся у матерей данного возраста. Исходная выборка (а это реальные данные по демографии США в 1920 г.) составляет 100000 матерей, а число их дочерей-116760. Чтобы уравновесить число дочерей (точнее, массу гистограммы) по левому плечу передвигается груз, равный массе гистограммы на правом плече. В приведенном примере равновесие было достигнуто, когда груз установлен на отметку 28,5 лет. Именно на этот возраст матери приходилось среднее для всей популяции рождение «среднего» ребенка (точнее, девочки) в США в 1920 г.
Поскольку скорость роста популяции находится в обратной зависимости от длительности поколения r = lnR 0 /T , очевидно, чем раньше происходит размножение организмов, тем больше скорость роста популяции. Поясним это на воображаемом примере двух человеческих популяций, растущих по экспоненциальному закону. Предположим, что в первой популяции у каждой женщины в среднем по 5 детей, причем первый ребенок появлялся у них в 18 лет, а затем каждый год рождалось по одному ребенку (последний в 22 года). Предположим, что во второй популяции у каждой женщины в среднем по 10 детей, но появлялись на свет они позже, когда матери было от 30 до 39 лет (как и в предыдущем случае, в год по ребенку). Сначала может показаться, что вторая популяция растет в два раза быстрее первой. Но не будем торопиться с выводами и подсчитаем специфическую скорость роста r. Предположим, что девочки составляют половину всех родившихся детей. Тогда число девочек, приходящихся на одну мать, будет в первом случае 2,5, а во втором - 5. Напомним, что отношение численности дочернего поколения к численности материнского поколения есть не что иное, как R 0 - чистая скорость. воспроизводства. Тогда для первой популяции R 0 = 2,5, а для второй R 0 = 5. Длительность поколения Т в первой популяции будет составлять 20 лет, а во второй - 34,5 года. Соответственно значение г для первой популяции будет r 1 = ln2.5/20 = 046 , а для второй r 2 = ln5/34.5 = 0,047.
Полученные величины практически одинаковы. Иными словами, женщины, родившие в возрасте от 18 до 22 лет 5 детей, вносят примерно такой же вклад в увеличение численности популяции, как и женщины, родившие в возрасте от 30 до 39 лет по 10 детей. Конечно, эти рассуждения справедливы только в том случае, если в обеих популяциях сохранится то же распределение рождаемости по возрастам, т. е. девочки, рожденные более молодыми матерями, сами начнут рожать с 18 лет, а те, что родились от матерей 30-39 лет, - только с 30 лет.
Из приведенного выше примера ясно, сколь важное значение g демографической политике любого государства имеют законы, ограничивающие минимальный допустимый возраст вступления в брак, а также другие мероприятия, поощряющие деторождение только в определенном возрасте.
У многих животных возраст достижения половозрелости и. возраст начала размножения могут сильно меняться в зависимости от конкретных условий существования. В менее благоприятных условиях размножение наступает позже, и, таким образом, скорость роста популяций снижается. Так, например, у полевки-экономки (Microtus oeconomus), численность которой регулярно колеблется, половозрелость может наступать на 20-25-й день в период нарастания численности или только на 9-11-й месяц в гиды пиковой численности и в период депрессии. У планктонного ветвистоусого рачка Diaphanosoma brachyurum, обычного в летнее время вида в планктоне озер умеренной зоны, при обилии пищи откладка самками партеногенетических яиц наблюдается на 5-6-й день после рождения, тогда как при нехватке пищи Размножение начинается только через 20-30 дней(при этом добивает до данного возраста только небольшая часть популяции).
Важнейшая особенность популяции, растущей по экспоненциальному закону, - это стабильная возрастная структура, т. е. постоянное соотношение численностей разных возрастных групп. Справедливо и обратное утверждение: если в популяции поддерживается постоянное соотношение разных возрастных групп (а это соотношение в свою очередь есть следствие не меняющихся во времени распределений l х и т х), то такая популяция растет экспоненциально. Конечно, в популяциях, растущих экспоненциально, но с разной скоростью, возрастная структура различна: чем быстрее растет численность популяции, тем больше доля молодых особей (рис. 26). Как частный случай экспоненциального роста можно рассматривать стационарную популяцию, не меняющую свою численность во времени (т. е. r = 0). В такой популяции также устанавливается стабильная возрастная структура.
Если наблюдать за возрастной структурой популяции какого-нибудь вида, продолжительность жизни которого по крайней мере несколько лет, можно заметить, как когорта молодых особей, появившихся на свет в благоприятный для размножения и (или) для выживания ранних стадий развития год, будучи многочисленнее других когорт, переходит из одной возрастной группы в другую. Такие «урожайные» поколения хорошо прослеживаются, например, в популяциях рыб.
Рис. 27. Возрастная структура населения Франции по данным на 1 января 1967 г. (слева - мужчины, справа - женщины). Пониженная численность мужского населения рождения 83-90-х гг. прошлого века - это результат массовой гибели во время первой мировой войны; «талия», приходящаяся на 1916 г. рождения, - это результат резкого снижения рождаемости в годы первой мировой войны; вторая «талия», приходящаяся на 40-е гг. рождения, - это результат снижения рождаемости в годы второй мировой войны; увеличение численности людей рождения 19-!6-1949 гг. - результат подъема рождаемости после массовой демобилизации. Непосредственные потери людей в годы второй мировой войны отражены прежде всего сокращением численности мужчин 1906-1926 гг. рождения (по Shrvock еt al., 1976; из Begon et al., 1986)
В возрастной структуре населения европейских стран мощный след оставили две мировые войны. Например, на гистограмме возрастного распределения населения Франции в 1967 г. (рис. 27) хорошо видны две «талии»: верхняя-это результат снижения рождаемости в годы первой мировой войны, а нижняя-результат снижения рождаемости в годы второй мировой войны. Каждый раз после окончания войны наблюдался подъем рождаемости.







