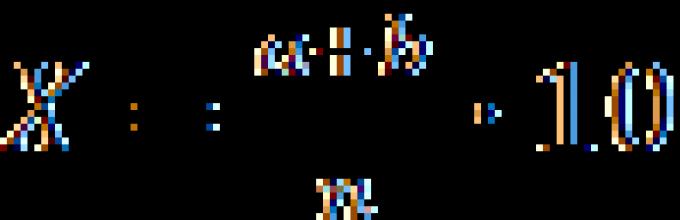
Хромосомный уровень организации наследственного материала. Хромосомы, как группы сцепления генов.
Из принципов генетического анализа вытекает, что независимое комбинирование признаков может осуществляться лишь при условии, что гены, определяющие эти признаки, находятся в разных парах хромосом. Следовательно, у каждого организма, число пар признаков, по которым наблюдается независимое наследование, ограничено числом пар хромосом. С другой стороны, очевидно, что число признаков и свойств организма, контролируемых генами, чрезвычайно велико, а число пар хромосом у каждого вида относительно мало и постоянно. Остается допустить, что в каждой хромосоме находится не один ген, а много. Если это так, то следует признать, что третье правило Менделя касается только распределения хромосом, а не генов, т.е. его действие ограничено. Анализ проявления третьего правила показал, что в некоторых случаях новые комбинации генов у гибридов совсем отсутствовали, т.е. наблюдалось полное сцепление между генами исходных форм и в фенотипе наблюдалось расщепление 1:1. В других случаях комбинация признаков отмечалась с меньшей частотой, чем ожидается при независимом наследовании.
В 1906 году У. Бетсон описал нарушение менделевского закона независимого наследования двух признаков. Возникли вопросы: почему не все признаки наследуются и как они наследуются, как расположены гены в хромосомах, каковы закономерности наследования генов, находящихся в одной хромосоме? На эти вопросы смогла ответить хромосомная теория наследственности, созданная Т. Морганом, в 1911 году.
Т. Морган, изучив все отклонения, предложил называть совместное наследование генов, ограничивающее их свободное комбинирование, сцеплением генов или сцепленным наследованием.
Закономерности полного и неполного сцепления. Группы сцепления у человека.
Исследования Т. Моргана и его школы показали, что в гомологичной паре хромосом регулярно происходит обмен генами. Процесс обмена идентичными участками гомологичных хромосом с содержащимися в них генами называют перекрестом хромосом или кроссинговером. Кроссинговер наблюдается в мейозе. Он обеспечивает новые сочетания генов, находящихся в гомологичных хромосомах. Явление кроссинговера, как и сцепление генов, характерно для животных, растений, микроорганизмов. Исключение составляют самцы дрозофилы и самки тутового шелкопряда. Кроссинговер обеспечивает рекомбинацию генов и тем самым значительно увеличивает роль комбинативной изменчивости в эволюции. О наличии кроссинговера можно судить на основе учета частоты возникновения организмов с новым сочетанием признаков. Явление кроссинговера было открыто Морганом на дрозофиле.
Запись генотипа дигетерозиготы при независимом наследовании:
А В
Запись генотипа дигетерозиготы при сцепленном наследовании:
Гаметы с хромосомами, претерпевшими кроссинговер, называют кроссоверными, а не претерпевшие – некроссоверными.
АВ, ав Ав, аВ
Некроссоверные гаметы. Кроссоверные гаметы.
Соответственно организмы, возникшие от сочетания кроссоверных гамет, называют кроссоверами или рекомбинантами, а возникшие от сочетания некроссоверных гамет – некроссоверами или нерекомбинантами .
Явление кроссинговера, как и сцепление генов, можно рассмотреть и в классическом опыте Т. Моргана при скрещивании дрозофил.
|
Признак |
P♀ BV x ♂ bv |
|
|
серый цвет тела |
||
|
черный цвет тела |
||
|
нормальные крылья |
||
|
рудиментарные крылья |
||
|
Анализирующее скрещивание 1. Полное сцепление генов. 2. Неполное сцепление генов. |
1. Полное сцепление P♀ bv x ♂ BV F 2 bv bv расщепление – 1:1 |
|
|
2. Неполное сцепление (кроссинговер) P:♀ BV x ♂ bv G: BV bv Bv bV bv некроссоверные кроссоверные F 2 BV bv Bv bV некроссоверов – 83% кроссоверов – 17% |
||
Для измерения расстояния между генами путем анализирующего скрещивания можно применять формулу:
![]() где:
где:
X – расстояние между генами в % кроссинговера или в морганидах;
а – количество особей 1-й кроссоверной группы;
в – количество особей 2-й кроссоверной группы;
n – общее количество гибридов в опыте;
100% – коэффициент для перевода в проценты.
На основании исследования сцепленного наследования Морган сформулировал тезис, вошедший в генетику под названием правило Моргана : гены, локализованные в одной хромосоме, наследуются сцеплено, причем сила сцепления зависит от расстояния между ними.
Сцепленные гены расположены в линейном порядке и частота кроссинговера между ними прямо пропорциональна расстоянию между ними. Однако, этот тезис характерен только для близко лежащих друг к другу генов. В случае же относительно удаленных генов наблюдается некоторое отклонение от такой зависимости.
Морган предложил выражать расстояние между генами в процентах кроссинговера между ними. Расстояние между генами также выражают в морганидах или сантиморганидах. Морганида – генетическое расстояние между генами, где происходит кроссинговер с частотой 1%.
По частоте кроссинговера между двумя генами можно судить об относительном расстоянии между ними. Так, если между генами А и В кроссинговер составляет 3%, а между генами В и С – 8% кроссинговера, то между А и С кроссинговер должен происходить с частотой либо 3+8=11%, либо 8-3=5%, в зависимости от того, в каком порядке эти гены расположены в хромосоме.
А ─ ─ ─ В ─ ─ ─ ─ ─ ─ ─ ─ С В ─ ─ ─ А ─ ─ ─ ─ ─ ─ ─ ─ С
Задача 1. Катаракта и полидактилия наследуются как доминантные аутосомные признаки. Женщина унаследовала катаракту от отца, полидактилию от матери. Гены сцеплены, расстояние между ними 3М. Каковы генотипы и фенотипы детей от брака этой женщины и мужчины нормального по этим признакам? Какова вероятность рождения здоровых детей?
|
катаракта |
P ♀ аВ х ♂ ав |
|
|
полидактилия |
Х = АВ = 3 Морг. |
|
|
P ♀ аВ х ♂ ав |
Ответ: вероятность рождения здорового ребенка – 1,5%, имеющих по 1 признаку – 48,5%, имеющих оба признака – 1,5% |
|
|
G: (аВ) (Ав) (ав) |
||
|
F1 аВ Ав ав АВ ав ав ав ав 48,5% 48,5% 1,5% 1,5% |
||
Генетическая карта хромосомы – это схема, отображающая порядок расположения генов на относительном расстоянии их друг от друга. О расстоянии между сцепленными генами судят по частоте кроссинговера между ними. Генетические карты всех хромосом составлены для наиболее изученных в генетическом отношении организмов: дрозофилы, кур, мышей, кукурузы, томатов, нейроспоры. Для человека также составлены генетические карты всех 23 хромосом.
После установления линейной дискретности хромосом возникла необходимость составления цитологических карт с целью сопоставления с генетическими, составленными на основе учета рекомбинаций.
Цитологическая карта – это карта хромосомы, на которой определяется расположение и относительное расстояние между генами в самой хромосоме. Построение их ведется на основе анализа хромосомных перестроек, дифференциальной окраски политенных хромосом, радиоактивных меток и др.
К настоящему времени, у ряда растений и животных построены и сопоставлены генетические и цитологические карты. Реальность этого сопоставления подтверждает правильность принципа о линейном расположении генов в хромосоме.
У человека можно назвать некоторые случаи сцепленного наследования.
Гены, контролирующие наследование групп крови по системе АВ0 и синдрома дефекта ногтей и коленной чашечки, наследуются сцепленно.
Сцеплены гены резус-фактора и овальной формы эритроцитов.
В третьей аутосоме расположены гены группы крови Лютеран и секреции антигенов А и В со слюной.
Гены полидактилии и катаракты наследуются сцепленно.
В Х-хромосоме расположены гены гемофилии и дальтонизма, а также гены цветовой слепоты и мышечной дистрофии Дюшена.
В 6 аутосоме находятся сублокусы А, В, С, D/DR системы HLA, контролирующих синтез антигенов гистосовместимости.
Наследование признаков Х-сцепленных и голандрических.
Признаки, контролируемые генами, расположенными в половых хромосомах, называются сцепленным с полом. У человека описано более 60 заболеваний, сцепленных с полом, большинство из которых наследуются рецессивно. Гены в половых хромосомах можно разделить на 3 группы:
Гены частично сцепленные с полом. Они расположены в парных сегментах Х и Y хромосом . К заболеваниям частично сцепленным с полом относят: геморрагический диатез, судорожные расстройства, пигментный ретинит, пигментную ксеродерму, общую цветовую слепоту.
Гены полностью сцепленные с полом. Они расположены в участке Х хромосомы , для которого нет гомологичного участка в Y хромосоме (гетерологическом). Эти гены контролируют заболевания: атрофия зрительного нерва, мышечная дистрофия Дюшена, дальтонизм, гемофилия, способность ощущать запах синильной кислоты.
Гены, расположенные в участке Y хромосомы , для которого нет гомологичного локуса в Х хромосоме, называются голандрическими . Они контролируют признаки: синдактилия, гипертрихоз ушной раковины.
Ген дальтонизма проявляется у 7% мужчин и у 0,5% женщин, но носительницами этого гена являются 13% женщин.
Сцепленное с полом наследование было описано Т. Морганом на примере наследования признака окраски глаз у дрозофилы.
Отмечено несколько закономерностей наследования сцепленных с полом признаков:
передаются крест на крест (от отца – дочери, от матери – сыну);
результаты прямого и обратного скрещиваний не совпадают;
у гетерогаметного пола признак проявляется в любом состоянии (доминантном или рецессивном).
Основные положения хромосомной теории наследственности.
Основные положения хромосомной теории наследственности можно сформулировать следующим образом:
Гены находятся в хромосомах. Каждый ген в хромосоме занимает определенный локус. Гены в хромосомах расположены линейно.
Каждая хромосома представляет группу сцепленных генов. Число групп сцепления у каждого вида равно числу пар хромосом.
Между гомологичными хромосомами происходит обмен аллельными генами – кроссинговер.
Расстояние между генами в хромосоме пропорционально проценту кроссинговера между ними. Зная расстояние между генами можно вычислить процентное соотношение генотипов у потомства.
В 1906 году У. Бэтсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве, гибриды всегда повторяли признаки родительских форм. Стало ясно, что не для всех признаков характерно независимое распределение в потомстве и свободное комбинирование.
Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно, каждая хромосома несет не один ген, а целую группу генов, отвечающих за развитие разных признаков. Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался Т. Морган . Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала плодовая мушка дрозофила.
Дрозофила каждые две недели при температуре 25 °С дает многочисленное потомство. Самец и самка внешне хорошо различимы — у самца брюшко меньше и темнее. Они имеют всего 8 хромосом в диплоидном наборе, достаточно легко размножаются в пробирках на недорогой питательной среде.
Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, — над геном недоразвитых). При проведении анализирующего скрещивания самки F 1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% — серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% — черные длиннокрылые и 8,5% — серые с зачаточными крыльями). Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме.

1 — некроссоверные гаметы; 2 — кроссоверные гаметы.
Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — АВ и аb , а отцовский — один тип — аb . Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип ААВВ и ааbb . Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Ааbb и ааВb . Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток — мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В , появляются гаметы Аb и аВ , и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1.
Группа сцепления — гены, локализованные в одной хромосоме и наследующиеся совместно. Количество групп сцепления соответствует гаплоидному набору хромосом.
Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот. Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным. Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Независимое наследование — наследование признаков, гены которых локализованы в разных парах гомологичных хромосом.
Некроссоверные гаметы — гаметы, в процессе образования которых кроссинговер не произошел.
Нерекомбинанты — гибридные особи, у которых такое же сочетание признаков, как и у родителей.
Рекомбинанты — гибридные особи, имеющие иное сочетание признаков, чем у родителей.
Расстояние между генами измеряется в морганидах — условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам.
У дигетерозигот доминантные гены могут располагаться или в одной хромосоме (цис-фаза ), или в разных (транс-фаза ).

1 — Механизм цис-фазы (некроссоверные гаметы); 2 — механизм транс-фазы (некроссоверные гаметы).
Результатом исследований Т. Моргана стало создание им хромосомной теории наследственности :
- гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов; набор генов каждой из негомологичных хромосом уникален;
- каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
- гены расположены в хромосомах в определенной линейной последовательности;
- гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
- сцепление генов может нарушаться в процессе кроссинговера, что приводит к образованию рекомбинантных хромосом; частота кроссинговера зависит от расстояния между генами: чем больше расстояние, тем больше величина кроссинговера;
- каждый вид имеет характерный только для него набор хромосом — кариотип.
Перейти к лекции №17 «Основные понятия генетики. Законы Менделя»
В 1906 году У. Бэтсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве, гибриды всегда повторяли признаки родительских форм. Стало ясно, что не для всех признаков характерно независимое распределение в потомстве и свободное комбинирование.
Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно, каждая хромосома несет не один ген, а целую группу генов, отвечающих за развитие разных признаков. Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался Т. Морган . Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала плодовая мушка дрозофила.
Дрозофила каждые две недели при температуре 25 °С дает многочисленное потомство. Самец и самка внешне хорошо различимы — у самца брюшко меньше и темнее. Они имеют всего 8 хромосом в диплоидном наборе, достаточно легко размножаются в пробирках на недорогой питательной среде.
Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, — над геном недоразвитых). При проведении анализирующего скрещивания самки F 1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% — серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% — черные длиннокрылые и 8,5% — серые с зачаточными крыльями). Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме.
1 — некроссоверные гаметы; 2 — кроссоверные гаметы.
Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — АВ и аb , а отцовский — один тип — аb . Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип ААВВ и ааbb . Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Ааbb и ааВb . Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток — мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В , появляются гаметы Аb и аВ , и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1.
Группа сцепления — гены, локализованные в одной хромосоме и наследующиеся совместно. Количество групп сцепления соответствует гаплоидному набору хромосом.
Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот. Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным. Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Независимое наследование — наследование признаков, гены которых локализованы в разных парах гомологичных хромосом.
Некроссоверные гаметы — гаметы, в процессе образования которых кроссинговер не произошел.
Нерекомбинанты — гибридные особи, у которых такое же сочетание признаков, как и у родителей.
Рекомбинанты — гибридные особи, имеющие иное сочетание признаков, чем у родителей.
Расстояние между генами измеряется в морганидах — условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам.
У дигетерозигот доминантные гены могут располагаться или в одной хромосоме (цис-фаза ), или в разных (транс-фаза ).
1 — Механизм цис-фазы (некроссоверные гаметы); 2 — механизм транс-фазы (некроссоверные гаметы).
Результатом исследований Т. Моргана стало создание им хромосомной теории наследственности :
- гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов; набор генов каждой из негомологичных хромосом уникален;
- каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
- гены расположены в хромосомах в определенной линейной последовательности;
- гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
- сцепление генов может нарушаться в процессе кроссинговера, что приводит к образованию рекомбинантных хромосом; частота кроссинговера зависит от расстояния между генами: чем больше расстояние, тем больше величина кроссинговера;
- каждый вид имеет характерный только для него набор хромосом — кариотип.
Перейти к лекции №17 «Основные понятия генетики. Законы Менделя»
Лекция №18. Сцепленное наследование
В 1906 году У. Бэтсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве, гибриды всегда повторяли признаки родительских форм. Стало ясно, что не для всех признаков характерно независимое распределение в потомстве и свободное комбинирование.
Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно, каждая хромосома несет не один ген, а целую группу генов, отвечающих за развитие разных признаков. Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался Т. Морган . Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала плодовая мушка дрозофила.
Дрозофила каждые две недели при температуре 25 °С дает многочисленное потомство. Самец и самка внешне хорошо различимы - у самца брюшко меньше и темнее. Они имеют всего 8 хромосом в диплоидном наборе, достаточно легко размножаются в пробирках на недорогой питательной среде.
Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, - над геном недоразвитых). При проведении анализирующего скрещивания самки F 1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% - серые длиннокрылые и 41,5% - черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% - черные длиннокрылые и 8,5% - серые с зачаточными крыльями). Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме.

1 - некроссоверные гаметы; 2 - кроссоверные гаметы.
Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов - АВ и аb, а отцовский - один тип - аb. Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип ААВВ и ааbb. Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Ааbb и ааВb. Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток - мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В, появляются гаметы Аb и аВ, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1.
Группа сцепления - гены, локализованные в одной хромосоме и наследующиеся совместно. Количество групп сцепления соответствует гаплоидному набору хромосом.
Сцепленное наследование - наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот. Полное сцепление - разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным. Неполное сцепление - разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Независимое наследование - наследование признаков, гены которых локализованы в разных парах гомологичных хромосом.
Некроссоверные гаметы - гаметы, в процессе образования которых кроссинговер не произошел.
Нерекомбинанты - гибридные особи, у которых такое же сочетание признаков, как и у родителей.
Рекомбинанты - гибридные особи, имеющие иное сочетание признаков, чем у родителей.
Расстояние между генами измеряется в морганидах - условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам.
У дигетерозигот доминантные гены могут располагаться или в одной хромосоме (цис-фаза ), или в разных (транс-фаза ).

1 - Механизм цис-фазы (некроссоверные гаметы); 2 - механизм транс-фазы (некроссоверные гаметы).
Результатом исследований Т. Моргана стало создание им хромосомной теории наследственности :
- гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов; набор генов каждой из негомологичных хромосом уникален;
- каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
- гены расположены в хромосомах в определенной линейной последовательности;
- гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
- сцепление генов может нарушаться в процессе кроссинговера, что приводит к образованию рекомбинантных хромосом; частота кроссинговера зависит от расстояния между генами: чем больше расстояние, тем больше величина кроссинговера;
- каждый вид имеет характерный только для него набор хромосом - кариотип.
Сцепленное наследование генов
В начале XX в., когда генетики стали проводить множество экспериментов по скрещиванию на самых различных объектах (кукуруза, томаты, мыши, мушки дрозофилы, куры и др.), обнаружилось, что не всегда проявляются закономерности, установленные Менделем . Например, не во всех парах аллелей наблюдается доминирование. Вместо него возникают промежуточные генотипы, в которых участвуют обе аллели. Обнаруживается также много пар генов, не подчиняющихся закону независимого наследования генов, особенно если пара аллельных генов находится в одной и той же хромосоме, т. е. гены как бы сцеплены друг с другом. Такие гены стали называть сцепленными .
Механизм наследования сцепленных генов, а также местоположение некоторых сцепленных генов установил американский генетик и эмбриолог Т. Морган . Он показал, что закон независимого наследования, сформулированный Менделем , действителен только в тех случаях, когда гены, несущие независимые признаки, локализованы в разных негомологичных хромосомах. Если же гены находятся в одной и той же хромосоме, то наследование признаков происходит совместно, т. е. сцепленно. Это явление стали называть сцепленным наследованием , а также законом сцепления или законом Моргана .
Закон сцепления гласит: сцепленные гены, расположеные в одной хромосоме, наследуются совместно (сцепленно) .
Примеров сцепленного наследования генов известно очень много. Например, у кукурузы окраска семян и характер их поверхности (гладкие или морщинистые), сцепленные между собой, наследуются совместно. У душистого горошка (Lathyrus odoratus) сцепленно наследуются окраска цветков и форма пыльцы.
Все гены одной хромосомы образуют единый комплекс - группу сцепления . Они обычно попадают в одну половую клетку - гамету и наследуются вместе.
Группа сцепления — все гены одной хромосомы. Число групп сцепления равно количеству хромосом в гаплоидном наборе. Например, у человека 46 хромосом — 23 группы сцепления, у гороха 14 хромосом — 7 групп сцепления, у плодовой мушки дрозофилы 8 хромосом — 4 группы сцепления.
Гены, входящие в группу сцепления, не подчиняются третьему закону Менделя о независимом наследовании. Однако полное сцепление генов встречается редко. Если гены располагаются близко друг к другу, то вероятность перекреста хромосом мала и они могут долго оставаться в одной хромосоме, а потому будут передаваться по наследству вместе. Если же расстояние между двумя генами на хромосоме велико, то существует большая доля вероятности, что они могут разойтись по разным гомологичным хромосомам. В этом случае гены подчиняются закону независимого наследования.
Неполное сцепление генов. При анализе наследования сцепленных генов было обнаружено, что иногда сцепление может нарушаться в результате кроссинговера, происходящего во время мейоза при образовании половых клеток.
Если место разрыва хромосом во время обмена участками расположено между генами А (а) и В(b), то появятся гаметы Ab и аВ , а в потомстве образуются четыре группы фенотипов, как при несцепленном наследовании генов. Отличие заключается в том, что численное соотношение фенотипов не будет соответствовать соотношению 1:1:1:1, как при дигибридном анализирующем скрещивании.
Чем дальше друг от друга находятся гены в хромосоме, тем выше вероятность перекрёста между ними, тем больше процент гамет с перекомбинированными генами, а следовательно, и больше процент особей, отличных от родителей. Такое явление называют неполным сцеплением генов .
На рисунке - Наследование при неполном сцеплении генов (на примере скрещивания двух линий дрозофил, где А - нормальные крылья, а - зачаточные крылья, В - серый цвет тела, в - черный цвет тела ).
Полное сцепление генов. Чем ближе друг к другу находятся гены в хромосоме, тем меньше вероятность перекрёста между ними. Если гены расположены очень близко друг к другу (рядом), то перекрёста между ними обычно не наблюдается. В этом случае говорят о полном сцеплении генов .







