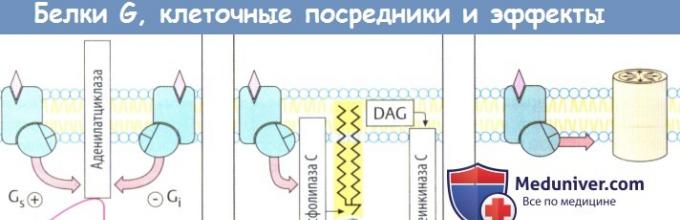
При передаче сигнала на рецепторы, сопряженные , участвует фактически такой же основной механизм. Связывание агониста рецептором приводит к изменению конформации белка рецептора. Это изменение передается на белок G: а-субъединица изменяет гуанозиндифосфат (ГДФ) на гуанозинтрифосфат (ГТФ), затем диссоциирует от двух других субъединиц, связывается с эффекторным белком и изменяет его функциональное состояние.
В принципе, β- и γ-субъединицы тоже способны взаимодействовать с эффекторными белками, а-субъединица медленно гидролизует связанный ГТФ до ГДФ . G a -ГДФ не обладает аффинитетом к эффекторному белку и вновь связывается с β- и γ-субъединицами. Белки G могут диффундировать вдоль мембраны; они не прикреплены к отдельным белкам рецепторов. Тем не менее существует связь между типами рецепторов и типами белка G.
Более того, α-субъединицы отдельных белков G различаются аффинитетом к разным эффекторным белкам, а также характером воздействия на них. G a -ГТФ белка G s стимулирует аденилатциклазу, в то время как G a -ГТФ белка G i является ее ингибитором.
Группа сопряженных с G-белком рецепторов включает мускариновые холинорецепторы, адренорецепторы норадреналина и адреналина, а также рецепторы дофамина, гистамина, серотонина, глутамата, ГАМК, морфина, простагландинов, лейкотриенов и многих других медиаторов и гормонов.
К основным эффекторным белкам у сопряженных с G-белком рецепторов относятся аденилатциклаза (АТФ => внутриклеточный посредник цАМФ), фосфолипаза С (фосфатидилинозитол => внутриклеточные посредники инозитолтрифосфата и диацилглицерола), а также белки ионного канала. Множество клеточных функций регулируется концентрацией клеточного циклического аденозинмонофосфата (цАМФ), поскольку цАМФ увеличивает активность протеинкиназы А, которая катализирует превращение фосфатных групп в функциональные белки.
Повышение уровня цАМФ приводит к снижению тонуса гладких мышц, увеличению сократимости сердечной мышцы, а также повышению гликогенолиза и липолиза. Фосфорилирование белков сердечных Са 2+ -каналов повышает вероятность открытия каналов во время деполяризации мембраны. Следует отметить, что цАМФ инактивируется фосфодиэстеразой. Ингибиторы данного фермента повышают внутриклеточную концентрацию цАМФ и действуют как адреналин.
Белок рецептора самостоятельно фосфорилируется с потерей свойства активировать связанный белок G. Это один из механизмов, который способствует снижению чувствительность клетки во время длительной стимуляции рецептора агонистом (десенситизация).
Активация фосфолипазы С приводит к гидролизу мембранного фосфолипида фосфатидилинозитол-4,5-дифосфата до инозитолтрифосфата (IР 3) и диацил-глицерола (DAG). Инозитолтрифосфат стимулирует высвобождение Са 2+ из органелл-депо, что приводит к сокращению гладкомышечных клеток, распаду гликогена или к активации экзоцитоза. Диацилглицерол стимулирует протеинкиназу С, которая фосфорилирует ряд ферментов, содержащих серин или треонин.
Некоторые белки G вызывают открытие белков каналов. Таким образом активируются К + -каналы (действие АХ на синусовый узел, действие опиоидов на передачу нервного импульса).

В покое на постсинаптической мембране находится комплекс рецептор‑G-белок c ГДФ (рис. 809251335, 809251304).
Использованные в рис. 809251335 обозначения фаз ГТФ-азного цикла G‑белков не являются обязательными. В литературе выделяют 5, 6 и более фаз. На рис. 809251304 использовано иное обозначения. Однако принципиальной разницы в представленных схемах нет. Рисунок 809251304 Вам при желании легче будет воспроизвести.

Рис. 809251304. ГТФ-азный цикла G‑белков. 1- фаза покоя; 2 - оединяется медиатора (лиганда) к рецептору; 3 - диссоциация G‑белка; 4 - активация эффектора; 5 - дефосфорилирование ГТФ. Красное окрашивание рецептора на рисунке символизирует активацию; R – рецептор; Е - эффектор; L – медиатор (лиганд, агонист).
Связывание медиатора (агониста) с рецептором приводит к белок-белковому взаимодействию между рецептором и G-белком и ускоряет диссоциацию ГДФ. В результате образуется короткоживущий комплекс агонист - рецептор - G-белок, не связанный ни с каким нуклеотидом.
Связывание с этим комплексом молекулы ГТФ снижает сродство рецептора к G‑белку, что приводит к диссоциации комплекса и высвобождению рецептора.
Потенциально рецептор может активировать большое количество молекул G-белка, обеспечивая, таким образом, высокий коэффициент усиления внеклеточного сигнала на данном этапе.
Активированная альфа-субъединица G‑белка диссоциирует от бета-гамма-субъединиц и вступает во взаимодействие с соответствующим эффектором, оказывая на него активирующее или ингибирующее воздействие.
Альфа-субъдиница с присоединенным с ней ГТФ способна взаимодействовать с эффектором в мембране - ферментами, такими, как аденилатциклаза, или, возможно, ионными каналами. Фермент может активироваться или ингибироваться, а ионный канал - открываться или закрываться. Конкретные примеры будут рассмотрены ниже и в последующих лекциях.
Взаимодействие с эффектором, однако, длится до тех пор, пока альфа-субъединица, являющаяся ГТФ-азой, удерживает ГТФ.
Вскоре присоединенный ГТФ гидролизуется до ГДФ. Когда это происходит, альфа‑субъединица снова меняет свою конформацию и теряет способность активировать эффектор. После этого альфа-ГДФ взаимодействует с бета-гамма-комплексом и снова образует тримерный комплекс, завершая, таким образом, цикл.
Пример прямой активация калиевого канала субъединицами G-белка показан на рис. 809251442.
Пример прямого ингибирования Са 2+ -канала субъединицами G-белка показан на рис. 809251458.
Активация пресинаптических адренорецепторов (ауторецепторов) приводит к диссоциации G-белка и последующему ингибированию (закрытию) кальциевых каналов, т.е. снижению уровня освобождение медиатора (норадреналина)
Пример метаботропной модуляции ионных каналов показан на рис. 709200023.
|
|

Рис. 709200023. Модуляция потенциал-зависимых кальциевых каналов норадреналином.
На GTP как молекулярный функциональный «выключатель» для регулировки клеточных процессов.
История
G-белки были обнаружены и исследованы Альфредом Гилманом и Мартином Родбеллом , которые получили за это открытие Нобелевскую премию по физиологии и медицине года .
Типы G-белков
G-белки делятся на две основных группы - гетеротримерные («большие») и «малые». Гетеротримерные G-белки - это белки с четвертичной структурой, состоящие из трёх субъединиц: альфа(α), бета (β) и гамма (γ). Малые G-белки - это белки из одной полипептидной цепи, они имеют молекулярную массу 20-25 кДа и относятся к суперсемейству Ras малых ГТФаз . Их единственная полипептидная цепь гомологична α-субъединице гетеротримерных G-белков. Обе группы G-белков участвуют во внутриклеточной сигнализации.
Гетеротримерные G-белки
У всех гетеротримерных G-белков сходный механизм активации: они активируются при взаимодействии со специфическими рецепторами, сопряженными с G-белками , при этом обменивая ГДФ на ГТФ и распадаясь на α- и βγ-субъединицы. α-субъединица, связанная с ГТФ, воздействует на следующее звено в цепи передачи сигнала. βγ-субъединица также может вызывать собственные эффекты. Инактивация G-белков происходит в результате медленного гидролиза ГТФ до ГДФ α-субъединицей, после чего происходит реассоциация (объединение) субъединиц.
Белки-помощники G-белков
В работе многих G-белков участвуют вспомогательные белки. GAPs (GTPase Activating Proteins, белки-активаторы ГТФазной активности) ускоряют гидролиз ГТФ, ускоряя инактивацию G-белков. Особенно важна функция GAPs для малых G-белков, так как альфа-субъединицы гетеротримерных G-белков часто сами обладают достаточной ГТФ-азной активностью. К GAP-белкам относятся белки семейства RGS.
GEFs (Guanine nucleotide Exchange Factors, факторы обмена гуаниловых нуклеотидов), ускоряют обмен ГДФ на ГТФ и таким образом активируют G-белки. Обычно для G-белка GEF-ом служит активированный лигандом рецептор, однако в некоторых случаях белки AGS (Activator of G-protein Signaling, активаторы передачи сигнала G-белками) могут активировать G-белок независимо от воздействия на него рецептора.
Напишите отзыв о статье "G-белки"
Примечания
Ссылки
- Eric R. Kandel, James H. Schwartz, Thomas M. Jessell. Principles of neural science. - New York: McGraw-Hill, 2000. - ISBN ISBN 0-8385-7701-6 .
- Lodish et al. 2000. Molecular Cell Biology 4th ed. W.H. Freeman and Company, New York.
- Voet, Donald and Judith G. Voet. 1995. Biochemistry 2nd ed. John Wilely & Sons, New York.
- Gilman A (1987). «G proteins: transducers of receptor-generated signals.». Annu Rev Biochem 56 : 615-49. PMID 3113327 .
- Neves S.R., Ram P.T., Iyengar R. (2002). «G Protein Pathways.». Science 296 : 1636-9. PMID: .
Внешние ссылки
Отрывок, характеризующий G-белки
– Да ведь прямо сказано: кто женится на разводной жене… – сказала старая княгиня.– Ah, maman, ne dites pas de betises. Vous ne comprenez rien. Dans ma position j"ai des devoirs, [Ах, маменька, не говорите глупостей. Вы ничего не понимаете. В моем положении есть обязанности.] – заговорилa Элен, переводя разговор на французский с русского языка, на котором ей всегда казалась какая то неясность в ее деле.
– Но, мой друг…
– Ah, maman, comment est ce que vous ne comprenez pas que le Saint Pere, qui a le droit de donner des dispenses… [Ах, маменька, как вы не понимаете, что святой отец, имеющий власть отпущений…]
В это время дама компаньонка, жившая у Элен, вошла к ней доложить, что его высочество в зале и желает ее видеть.
– Non, dites lui que je ne veux pas le voir, que je suis furieuse contre lui, parce qu"il m"a manque parole. [Нет, скажите ему, что я не хочу его видеть, что я взбешена против него, потому что он мне не сдержал слова.]
– Comtesse a tout peche misericorde, [Графиня, милосердие всякому греху.] – сказал, входя, молодой белокурый человек с длинным лицом и носом.
Старая княгиня почтительно встала и присела. Вошедший молодой человек не обратил на нее внимания. Княгиня кивнула головой дочери и поплыла к двери.
«Нет, она права, – думала старая княгиня, все убеждения которой разрушились пред появлением его высочества. – Она права; но как это мы в нашу невозвратную молодость не знали этого? А это так было просто», – думала, садясь в карету, старая княгиня.
В начале августа дело Элен совершенно определилось, и она написала своему мужу (который ее очень любил, как она думала) письмо, в котором извещала его о своем намерении выйти замуж за NN и о том, что она вступила в единую истинную религию и что она просит его исполнить все те необходимые для развода формальности, о которых передаст ему податель сего письма.
«Sur ce je prie Dieu, mon ami, de vous avoir sous sa sainte et puissante garde. Votre amie Helene».
[«Затем молю бога, да будете вы, мой друг, под святым сильным его покровом. Друг ваш Елена»]
Это письмо было привезено в дом Пьера в то время, как он находился на Бородинском поле.
Во второй раз, уже в конце Бородинского сражения, сбежав с батареи Раевского, Пьер с толпами солдат направился по оврагу к Князькову, дошел до перевязочного пункта и, увидав кровь и услыхав крики и стоны, поспешно пошел дальше, замешавшись в толпы солдат.
Одно, чего желал теперь Пьер всеми силами своей души, было то, чтобы выйти поскорее из тех страшных впечатлений, в которых он жил этот день, вернуться к обычным условиям жизни и заснуть спокойно в комнате на своей постели. Только в обычных условиях жизни он чувствовал, что будет в состоянии понять самого себя и все то, что он видел и испытал. Но этих обычных условий жизни нигде не было.
Хотя ядра и пули не свистали здесь по дороге, по которой он шел, но со всех сторон было то же, что было там, на поле сражения. Те же были страдающие, измученные и иногда странно равнодушные лица, та же кровь, те же солдатские шинели, те же звуки стрельбы, хотя и отдаленной, но все еще наводящей ужас; кроме того, была духота и пыль.
Пройдя версты три по большой Можайской дороге, Пьер сел на краю ее.
Лекарства, действующие на рецепторы , вызывают широкий спектр тканевых и системных ответов по двум причинам. Во-первых, на разных тканях находятся различные рецепторы. Во-вторых, различным типам рецепторов присуща разная структура и, соответственно, функция, поэтому клеточный ответ на активацию рецептора (трансдукция) значительно варьирует в зависимости от структуры рецептора. Выделяют четыре типа рецепторов, которые отличаются кинетическими характеристиками:
G-белок-связанные рецепторы;
ДНК-связанные рецепторы;
рецепторы, обладающие тирозинкиназной активностью (тирозинкиназные рецепторы);
Рецептор-связанные каналы (РСК).
Тирозинкиназные рецепторы и РСК отличаются от остальных тем, что для проявления клеточного ответа они не требуют клеточной трансдукции компонентов при активации агонистом, поэтому молекула рецептора играет большую роль, чем молекулярное звено между лекарством и трансдукцией. Тирозинкиназный рецептор - в действительности мембраносвязанный фермент, который «включается» агонистом. РСК - специфический ионный канал, который структурно отличается от потенциалзависимых каналов, регулируемых лекарством или нейротрансмиттером, тем, что он присоединяется к высокоспецифическому лиганд-связывающему центру, а не к специфическому потенциалчувствительному участку. В РСК лиганд-связывающий центр и канал являются функционально различными участками одной молекулы. РСК также называют лиганд-зависимым ионным каналом. Приведенная классификация удобна, поскольку рассматривает тирозинкиназные структуры и РСК как рецепторы.
G-белок-связанные рецепторы
G-белки являются компонентами трансдукции. Рецепторы, связанные с G-белками, локализованы на клеточной мембране и состоят из 7 трансмембранных спиралей (I-VII).
В отсутствие агониста рецептор связан с G-белком, что удерживает рецептор в неактивной конформации. Сам по себе - это комплекс трех субъединиц (а, бета и у). Когда рецептор находится в состоянии покоя, три субъединицы G-белка и гуанозиндифосфат (ГДФ) прочно связаны с альфа-субъединицей G-белка. Связывание агониста приводит к конформационным изменениям рецептора, что, в свою очередь, вызывает конформационное изменение G-белка, ведущее к диссоциации ГДФ от альфа-субъединицы. Это инициирует последовательность реакций и способствует трансдукции G-белок-связанного рецептора, которая описывается далее.
Из компонентов трансдукции , непосредственно связанных с G-белками, наиболее распространенным в организме является аденилилциклаза. Циклический нуклеотид цАМФ синтезируется из аденозинтрифосфата (АТФ) ферментом аденилилциклазой. цАМФ обладает различными биологическими эффектами.
цАМФ оказывает действие на энергетический обмен, клеточную дифференцировку, функцию ионных каналов и сократительные белки
цАМФ фосфорилирует внутриклеточные белки (чаще ферменты) через воздействие цАМФ-зависимых протеинкиназ. Эти протеинкиназы активируют цАМФ и фосфорилируют аминокислоты серии и треонин, используя АТФ как источник фосфора. Результат фосфорилирования состоит в:
Активации гормон-чувствительных липаз;
инактивации гликогенсинтазы;
активации фосфорилазкиназы и превращении неактивной фосфорилазы в активную, что приводит к ускорению липолиза, снижению синтеза и ускорению распада гликогена;
активации Са2+-каналов L-типа и саркоплазматического ретикулума в сердечных клетках за счет фосфорилирования, что увеличивает выход кальция.
Важнейшим внутриклеточным компонентом сигнальных каскадов являются G-белки. В настоящее время известно около 20 различных G-белков. Так, например, Gs и Gi стимулируют и ингибируют аденилатциклазу, соответственно; Gq активирует фосфолипазу С. Среди G-белков сенсорных клеток можно отметить: фоторецепторные - Gt (трансдуцин), обонятельные - Golf и вкусовые - Gg.
По своему строению G-белки представляют собой гетеротримеры, состоящие из трех типов субъединиц: a (альфа), b (бетта) и g (гамма), однако в нативных условиях бетта и гамма субъединицы функционируют как единый комплекс. Общей структурной особенностью G-белков является наличие семи трансмембранных альфа-спиралей. Важнейшей характеристикой G-белков является присутствие на их a-субъединице центра связывания гуаниловых нуклеотидов: GDP(гуанизидиндифосфат) и GTP (гуанизидинтрифосфат). Если с G-белком связан GTP, то это соответствует его активированному состоянию (G-GTP) или, иначе, G-белок находится в активированном положении. Если в нуклеотидсвязывающем центре присутствует GDP, то эта форма (G-GDP) соответствует "выключенному" состоянию. Ключевым моментом передачи сигнала от рецептора (на который подействовал первичный сигнал) к G-белку является катализ активированным рецептором обмена GDP, связанного с G-белком, на присутствующий в среде GTP (GDP / GTP-обмен на G-белке).
Трансмембранные рецепторы обеспечивают основные жизненно важные функции клетки: сигнальную, транспортную, защитную. Изучение механизма действия различных биологически активных соединений, в том числе противовирусных и противобактериальных показало, что наиболее специфичными мишенями, как для лекарственных, так и для токсических соединений (ядов) являются клеточные рецепторы человека и патогенных микроорганизмов. Значительную часть трансмембранных рецепторов составляют G-белок сопряженные рецепторы (GPCR), около половины всех известных в настоящее время лекарственных препаратов действуют именно на GPCR. Из всех видов поверхностных клеточных рецепторов GPCR наиболее универсальны. Эти рецепторы связывают широкий круг молекул, от, небольших по размеру нейромедиаторов, до крупных белков. GPCR вовлечены практически во все жизненно важные процессы.
Разнообразие сигналов, передаваемых GPCR, обеспечивается функциональным сопряжением разных GPCR между собой. Таким образом, очевидно, что наиболее универсальный механизм влияния токсичных и лекарственных соединений на клетку реализуется через воздействие на рецепторный аппарат клетки, путем изменения их конформации или основных характеристик связи лиганд-рецептор, их специфичности и обратимости.







