Ядро клетки - это одна из основных составных частей всех растительных и животных клеток, неразрывно связанная с обменом, передачей наследственной информации и др.
Форма ядра клетки варьирует в зависимости от типа клетки. Имеются овальные, шаровидные и неправильной формы - подковообразные или многолопастные ядро клетки (у лейкоцитов), четковидные ядра клетки (у некоторых инфузорий), разветвленные ядра клетки (в железистых клетках насекомых) и др. Величина ядра клетки различна, но обычно связана с объемом цитоплазмы. Нарушение этого соотношения в процессе роста клетки приводит к клеточному делению. Количество ядер клетки также неодинаково - большинство клеток имеет одно ядро, хотя встречаются двуядерные и многоядерные клетки (например, некоторые клетки печени и костного мозга). Положение ядра в клетке является характерным для клеток каждого типа. В зародышевых клетках ядро обычно находится в центре клетки, но может смещаться по мере развития клетки и образования в цитоплазме специализированных участков или отложения в ней резервных веществ.
В ядре клетки различают основные структуры: 1) ядерную оболочку (ядерную мембрану), через поры которой осуществляется обмен между ядром клетки и цитоплазмой [имеются данные, указывающие на то, что ядерная мембрана (состоящая из двух слоев) без перерыва переходит в мембраны эндоплазматической сети (см. ) и комплекса Гольджи]; 2) ядерный сок, или кариоплазму,- полужидкую, слабо окрашиваемую плазматическую массу, заполняющую все ядра клетки и содержащую в себе остальные компоненты ядра; 3) (см.), которые в неделящемся ядре видны только с помощью специальных методов микроскопии (на окрашенном срезе неделящейся клетки хромосомы обычно имеют вид неправильной сети из темных тяжей и зернышек, в совокупности называемых ); 4) одно или несколько сферических телец - ядрышек, являющихся специализированной частью ядра клетки и связанных с синтезом рибонуклеиновой кислоты и белков.
Ядро клетки обладает сложной химической организацией, в которой важнейшую роль играют нуклеопротеиды - продукт соединения с белками. В жизни клетки имеются два основных периода: интерфазный, или метаболический, и митотический, или период деления. Оба периода характеризуются главным образом изменениями в строении ядра клетки. В интерфазе ядро клетки находится в покоящемся состоянии и участвует в синтезе белков, регуляции формообразования, процессах секреции и других жизненных отправлениях клетки. В период деления в ядре клетки происходят изменения, приводящие к перераспределению хромосом и образованию дочерних ядер клетки; наследственная информация передается, таким образом, через ядерные структуры новому поколению клеток.
Ядра клетки размножаются только делением, при этом в большинстве случаев делятся и сами клетки. Обычно различают: прямое деление ядра клетки путем перешнуровки - амитоз и самый распространенный способ деления ядер клетки- типичное непрямое деление, или митоз (см.).
Действие ионизирующей радиации и некоторых других факторов способно изменять заключенную в ядре клетки генетическую информацию, приводя к различным изменениям ядерного аппарата, что иногда может приводить к гибели самих клеток или служить причиной наследственных аномалий у потомства (см. Наследственность), Поэтому изучение структуры и функций ядра клетки, особенно связей между хромосомными соотношениями и наследованием признаков, которыми занимается цитогенетика, имеет существенное практическое значение для медицины (см. ).
См. также Клетка.
Ядро клетки - важнейшая составная часть всех растительных и животных клеток.
Клетка, лишенная ядра или с поврежденным ядром, не способна нормально выполнять свои функции. Ядро клетки, точнее, организованная в его хромосомах (см.) дезоксирибонуклеиновая кислота (ДНК),- носитель наследственной информации, определяющей все особенности клетки, тканей и целого организма, его онтогенез и свойственные организму нормы реагирования на воздействия среды. Заключенная в ядре наследственная информация закодирована в составляющих хромосомы молекулах ДНК последовательностью четырех азотистых оснований: аденина, тимина, гуанина и цитозина. Эта последовательность является матрицей, определяющей структуру синтезируемых в клетке белков.
Даже самые незначительные нарушения структуры ядра клетки ведут к необратимым изменениям свойств клетки или к ее гибели. Опасность ионизирующих излучений и многих химических веществ для наследственности (см.) и для нормального развития плода имеет в своей основе повреждения ядер в половых клетках взрослого организма или в соматических клетках развивающегося эмбриона. В основе преобразования нормальной клетки в злокачественную также лежат определенные нарушения структуры ядра клетки.
Размеры и форма ядра клетки и соотношение его объема и объема всей клетки характерны для различных тканей. Одним из главных признаков, отличающих элементы белой и красной крови, являются форма и размер их ядер. Ядра лейкоцитов могут быть неправильной формы: изогнуто-колбасовидной, лапчатой или четковидной; в последнем случае каждый участок ядра соединен с соседним тонкой перемычкой. В зрелых мужских половых клетках (сперматозоидах) ядро клетки составляет подавляющую часть всего объема клетки.
Зрелые эритроциты (см.) человека и млекопитающих не имеют ядра, так как они теряют его в процессе дифференцировки. Они имеют ограниченный срок жизни и не способны размножаться. В клетках бактерий и сине-зеленых водорослей отсутствует резко очерченное ядро. Однако в них содержатся все характерные для ядра клетки химические вещества, распределяющиеся при делении по дочерним клеткам с такой же правильностью, как и в клетках высших многоклеточных организмов. У вирусов и фагов ядро представлено единственной молекулой ДНК.
При рассмотрении покоящейся (неделящейся) клетки в световом микроскопе ядро клетки может иметь вид бесструктурного пузырька с одним или несколькими ядрышками. Ядро клетки хорошо красится специальными ядерными красками (гематоксилин, метиленовый синий, сафранин и др.), которые обычно используют в лабораторной практике. При помощи фазово-контрастного устройства ядро клетки можно исследовать и прижизненно. В последние годы для изучения процессов, протекающих в ядре клетки, широко используют микрокинематографию, меченые атомы С14 и Н3 (ауторадиография) и микроспектрофотометрию. Последний метод особенно успешно применяют для изучения количественных изменений ДНК в ядре в процессе жизненного цикла клетки. Электронный микроскоп позволяет выявить детали тонкой структуры ядра покоящейся клетки, необнаруживаемые в оптическом микроскопе (рис. 1).
Рис. 1. Современная схема строения клетки, основанная на наблюдениях в электронном микроскопе: 1 - цитоплазма; 2 - аппарат Гольджи; 3 - центросомы; 4 - эндоплазматический ретикулум; 5 - митохондрии; 6 - оболочка клетки; 7 - оболочка ядра; 8 - ядрышко; 9 - ядро.
При делении клеток - кариокинезе или митозе (см.) - ядро клетки претерпевает ряд сложных преобразований (рис. 2), во время которых становятся отчетливо видимыми его хромосомы. Перед делением клетки каждая хромосома ядра синтезирует из веществ, присутствующих в ядерном соке, себе подобную, после чего материнская и дочерняя хромосомы расходятся к противоположным полюсам делящейся клетки. В результате каждая дочерняя клетка получает такой же хромосомный набор, какой был у материнской клетки, а вместе с ним и заключенную в нем наследственную информацию. Митоз обеспечивает идеально правильное разделение всех хромосом ядра на две равнозначные части.
Митоз и мейоз (см.) являются важнейшими механизмами, обеспечивающими закономерности явлений наследственности. У некоторых простейших организмов, а также в патологических случаях в клетках млекопитающих и человека ядра клетки делятся путем простой перетяжки, или амитоза. В последние годы показано, что и при амитозе происходят процессы, обеспечивающие разделение ядра клетки на две равнозначные части.
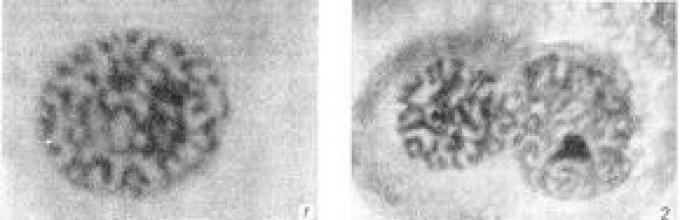
Набор хромосом в ядре клетки особи называют кариотипом (см.). Кариотип во всех клетках данной особи, как правило, одинаков. Многие врожденные аномалии и уродства (синдромы Дауна, Клайнфелтера, Тернера-Шерешевского и др.) обусловлены различными нарушениями кариотипа, возникшими либо на ранних стадиях эмбриогенеза, либо при созревании половой клетки, из которой возникла аномальная особь. Аномалии развития, связанные с видимыми нарушениями хромосомных структур ядра клетки, называют хромосомными болезнями (см. Наследственные болезни). Различные повреждения хромосом могут быть вызваны действием физических или химических мутагенов (рис. 3). В настоящее время методы, позволяющие быстро и точно устанавливать кариотип человека, используют для ранней диагностики хромосомных болезней и для уточнения этиологии некоторых заболеваний.

Рис. 2. Стадии митоза в клетках культуры ткани человека (перевиваемый штамм НЕр-2): 1 - ранняя профаза; 2 - поздняя профаза (исчезновение ядерной оболочки); 3 - метафаза (стадия материнской звезды), вид сверху; 4 - метафаза, вид сбоку; 5 - анафаза, начало расхождения хромосом; 6 - анафаза, хромосомы разошлись; 7 - телофаза, стадия дочерних клубков; 8 - телофаза и разделение клеточного тела.

Рис. 3. Повреждения хромосом, вызываемые ионизирующей радиацией и химическими мутагенами: 1 - нормальная телофаза; 2-4 - телофазы с мостами и фрагментами в эмбриональных фибробластах человека, облученных рентгеновыми лучами в дозе 10 р; 5 и 6 - то же в кроветворных клетках морской свинки; 7 - хромосомный мост в эпителии роговицы мыши, облученной дозой в 25 р; 8 - фрагментация хромосом в эмбриональных фибробластах человека в результате воздействия нитрозоэтилмочевиной.
Важный органоид ядра клетки - ядрышко - является продуктом жизнедеятельности хромосом. Оно продуцирует рибонуклеиновую кислоту (РНК), являющуюся обязательным промежуточным звеном в синтезе белка, вырабатываемого каждой клеткой.
Ядро клетки отделено от окружающей цитоплазмы (см.) оболочкой, толщина которой 60-70 Å.
Через поры в оболочке вещества, синтезируемые в ядре, поступают в цитоплазму. Пространство между оболочкой ядра и всеми его органоидами заполнено кариоплазмой, состоящей из основных и кислых белков, ферментов, нуклеотидов, неорганических солей и других низкомолекулярных соединений, необходимых для синтеза дочерних хромосом при делении ядра клетки.
Ядро, его строение и биологическая роль.
Ядро состоит из 1)поверхн аппарата ядра (в нем выдлел: 2 мембраны, перинуклеарн пространств, поровые комплексы, ламину.) 2) кариоплазмы (нуклеоплазмы) 3) хроматина (в нём эухроматин и гетерохроматин) 4) ядрышка (грануляр и фибриляр компонент.)
Ядро – это структура клетки которая выполняет функцию хранения и передачи инф, а так же регулирует все жизненные процессы клетки. Ядро несёт в себе генетическую (наследственную) инф в виде ДНК. Ядра обычно имеют шаровидную или яйцевидную форму. Я. окружено ядерн оболочкой. Ядерная оболочка пронизана ядерными порами. Через них ядро обменивается веществами с цитоплазмой(внутр средой клетки). Наружная мембрана переходит в эндоплпзматич ретикулум и может быть усеяна рибосомами. Отношение размеров ядра и клетки зависит от функциональной активности клетки. Большинство клеток одноядерные. Двуядерными могут быть кардиомиоциты. Всегда двуядерны инфузории. В них характерен ядерный дуализм.(то есть ядра различ по строению и финкциям). Малое ядро (генеративное) – диплойдное. Оно обеспечивает только половой процесс у инфузорий. Большое (вегетативное) ядро полиплойдное. Оно регулирует все остальные жизненные процессы. Многоядерными бывают клетки некоторых простейших и клетки скелетной мускулатуры.
П.А.Я. или кариотека ) имеет микроскопическую толщину и поэтому виден в световой микроскоп. Поверхностный аппарат ядра включает:
а)ядерную оболочку, или кариолемму;. б)паровые комплексы; в)периферическую плотную пластинку (ППП), или ламину.
(1) Ядерная оболочка (кариолемма). состоит из 2 мембран - наружной и внутренней, разделённых перинукляеарным пространством. Обе мембраны имеют такое же жидкосто-мозаичное строение, как и плазматическая мембрана, и различаются по набору белков. Среди этих белков имеются ферменты, переносчики и рецепторы. Наружная ядерная мембрана является продолжением мембран грЭПС и может быть усеяна рибосомами, на которых идёт синтез белка. Со стороны цитоплазмы наружная мембрана окружена сетью промежуточных (ви-ментиновых) фипаментов. Между наружной и внутренней мембранами находится перинуклеарное пространство -полость шириной 15-40 нм, содержимое которого сообщается с полостями каналов ЭПС. По составу перинуклеарное пространство близко к гиалоплазме и может содержать синтезированные рибосомами белки. Главная функция кариолеммы - изоляция гиалоплазмы от кариоплазмы. Специальные белки ядерных мембран, расположенные в области ядерных пор, осуществляют транспортную функцию. Ядерная оболочка пронизана ядерными порами, через которые осуществляется связь кариоплазмы и гиалоплазмы. Для регуляции такой связи в порах находятся (2) поровые комплексы. Они занимают 3-35% поверхности ядерной оболочки. Число ядерных пор с поровыми комплексами является изменчивой величиной и зависит от активности ядра. В области ядерных пор наружная и внутренняя ядерные мембраны сливаются. Совокупность структур, связанных с ядерной порой, называется комплексом ядерной поры. Типичный поровый комплекс представляет собой сложную белковую структуру - содержит более 1000 молекул белка. В центре поры расположена центральная белковая глобула (гранула), от которой по радиусу отходят тонкие фибриллы к периферическим белковым глобулам, образуя диафрагму поры. По периферии ядерной поры находятся две параллельные кольцевые структуры диаметром 80-120 нм (по одному с каждой поверхности кариолеммы), каждое из которых образовано 8 белковыми гранулами (глобулами).
Белковые глобулы перового комплекса подразделяются на центральные и периферические . С помощью периферических глобул осуществляется транспорт макромолекул из ядра в гиалоплазму. (фиксируются в мембране специальным интегральным белком. От этих гранул к центру сходятся белковые фибриллы, формирующие перегородку - диафрагму поры)
В нем участвуют специальные белки периферических глобул - нуклеопорины. В периферических глобулах имеется особый белок - переносчик молекул т-РНК
Центральная глобула специализируется на транспорте и-РНК из ядра в гиалопдазму. В её составе имеются ферменты, участвующее в химической модификации иРНК - ее процессинге.
Гранулы поровых комплексов структурно связаны с белками ядерной ламины, которая участвует в их организации
Функции комплекса ядерной поры:
1. Обеспечение регуляции избирательного транспорта в-в между цитоплазмой и ядром.
2. Активный перенос в ядро белков
3. Перенос в цитоплазму субъединиц рибосом
(3) ППП или ламина
слой толщиной 80-300 нм. прилегает изнутри к внутренней ядерной мембране. Внутренняя ядерная мембрана гладкая, ее интегральные белки связаны с ламиной (периферической плотной пластинкой). Ламина состоит из специальных переплетенных белков-ламинов, образующих периферический кариоскелет. Белки-ламины относятся к классу промежуточных филаментов (скелетных фибрилл). У млекопитающих известно 4 вида этих белков - это ломимы А, В, В 2 и С. Эти белки поступают в ядро из цитоплазмы. Ламины разных видов взаимодействуют между сбой и образуют белковую сеть под внутренней мембраной ядерной оболочки. С помощью ламинов «В» ППП соединяется со спец интеграл белкомядерн оболочки. С ППП взаимодействуют и белки приферич голобул «внутр кольца» порового комплекса. К ламину «А» присоед теломерн участки хромосом.
Функции ламины: 1) поддерд форму ядра. (даже есл бое мембраны разруш, то ядро за счет ламины сохр свою форму и поровые комп-сы ост на своём месте.
2) служит компонентом кариоскелета
3) участв в сборке ядерн оболочки (формирование кариоллемы) при делен клетки.
4) в интерфазном ядре к ламине прикрепл хроматин. таким образом ламина обеспеч функцию фиксации хроматина в ядре (обеспеч упорядочн укладку хроматина, участвует в пространственной организации хроматина в интерфазном ядре). Ламин «А» взаимодейств с теломерными участками хромосом.
5) обеспеч структур организацию поровых комплексов.
импорт и экспор белков.
В ядро через ядерные поры поступают: синтезированные цитоплазматическими рибосомами белки-ферменты, которые участвуют в процессах репликации и репарации (восстановления повреждений в ДНК); белки-ферменты, участвующие в процессе транскрипции; белки-репрессоры, которые регулируют процесс транскрипции; белки-гистоны.(которые связаны с молекулой ДНК и образуют хроматин); белки, входящие в состав субъединиц рибосом: белки ядерного матрикса, образующие кариоскелет; нуклеотиды; ионы минеральных солей, в частности, ионы Са и Mg .
Из ядра в цитоплазму выходят и-РНК. т-РНК и субъединицы рибосом, которые представляют собой рибонуклеопротеидные частицы (р-РНК, связанные с белками).
5. Химический состав и структурная организация хроматина. уровни компактизации. хромосомы чел их строен и классификация.
В ядре клеток мелкие зернышки и глыбки материала, окрашиваются основными красителями.
Хроматин представляет собой дезоксирибонуклеопротеид (ДНП) и состоит из ДНК, соединённой с белка-ми-гистонами или негистоновыми белками. Гистоны и ДНК объединены в структуры, которые называются нуклеосомами. Хроматин соответствует хромосомам, которые в интерфазном ядре представлены длинными перекрученными нитями и неразличимы как индивидуальные структуры. Выраженность спирализации каждой из хромосом неодинакова по их длине. Реализацию генетической информации осуществляют деспирализованные участки хромосом.
классификация хроматина:
1) эухроматин (активный деспирализованный. на нем происход считывание инф (транскрипция). в ядре выявляется как более светлые участки ближе к центру ядра) Предполагается, что в нем сосредоточена та ДНК, которая в интерфазе генетически активна. Эухроматин соответствует сегментам хромосом, которые деспирализованы и открыты для транскрипции.
2) гетерохроматин (нерабочий спирализованный, конденсированный, более компактный В ядре выявляется в виде глыбок на периферии.) делится на: конститутивный (всегда неактивен, никогда не переходит в эухроматин) и Факультативный (при определён условиях или на определен стадиях иммунного цикла может переходить в эухроматин). располагается ближе к оболочке ядра, более компактный. Примером скопления факульт гетерохроматина является тельце Барра - инактивированная Х-хромосома у самок млекопитающих, которая в интерфазе плотно скручена и неактивна.
Таким образом, по морфологическим признакам ядра (по соотношению содержания эу- и гетерохроматина) можно оценить активность процессов транскрипции, а, следовательно, синтетической функции клетки.
Хроматин и хромосомы представляют собой дезоксирибонуклеопротеиды (ДНП), но хроматин - это раскрученное, а хромосомы - скрученное состояние. Хромосом в интерфазном ядре нет, хромосомы появляются при разрушении ядерной оболочки (во время деления).
Строение хромосом:
хромосомы - наиболее упакованное состояние хроматина.
В хромосомах различают первичную перетяжку (центромеру), разделяющую хромосому на два плеча. Первичная перетяжка - наименее спирализованная часть хромосомы, к ней во время деления клетки присоединяются нити веретена деления. На некоторых хромосомах есть глубокие вторичные перетяжки, отделяющие небольшие участки хромосом, называемые спутниками. В области вторичных перетяжек находятся гены, кодирующие информацию об р-РНК, поэтому вторичные перетяжки хромосом называются ядрышковыми организаторами.
В зависимости от места расположения центромеры различают три типа хромосом:
1) метацентрические (имеют плечи равной или почтиравной величины);
2) субметацентрические (имеют плечи неравной величины);
3) акроцентрические (имеют палочковидную форму с коротким, почти незаметным вторым плечом);
Концы плеч хромосом называются теломерами
Уровни компаюпизации хроматина:
1. Нуклеосомный - Два с половиной витка двойной спирали ДНК (в 146-200 пар нуклеотидов) наматываются снаружи на белковый кор, образуя нуклеосому. Каждый гистон представлен двумя молекулами. ДНК наматывается на кор снаружи, образуя два с половиной витка. Участок ДНК между нуклеосомами называется линкером и имеет протяжбенность 50-60 пар нуклеотидов. Толщина нуклеосомной нити составляет 8-11 нм.
2. Нуклеомерный. Нуклеосомная структура закручивается, образуя суперспираль. В её образовании принимает участие ещё один гистоновый белок HI, лежащий между нуклеосомами и связанный с линкером. К каждому линкеру присоединяется 1 молекула гистона HI. Молекулы HI в комплексе с линкерами взаимодействуют между собой и вызывают суперспирализацию нуклеосомной фибриллы.
В результате образуется хроматиновая фибрилла, толщина которой составляет 30 нм (ДНК компактизирована в 40 раз). Суперспирализация происходит двумя способами. 1) нуклеосомная фибрилла может образовывать спираль второго порядка, которая имеет форму соленоида; 2) 8-10 нуклеосом образуют крупную компактную структуру - нуклеомеру. Этот уровень не допускает синтеза РНК с нуклеомерной ДНК (транскрипция не происходит).
3. Хромомерный (петельная структура). Хроматиновая фибрилла образует петли, которые сцепляются между собой с помощью особых негистоновых белков, либо петельные центры - хромомеры. Толщина 300 нм.
4. Хромонемный - образуется в результате сближения хромомеров по длине. Хромонема содержит одну гигантскую молекулу ДНК в комплексе с белками, т.е. фибриллу дезокси-рибонуклеопротеина - ДНП (400 нм).
5. Хроматидный - хромонема складывается несколько раз, образуя тело хроматиды (700 нм). После репликации ДНК хромосома содержит 2 хроматиды.
6. Хромосомный (1400 нм). Состоит из двух хроматид. Хроматиды соединены центромерой. При делении клетки хроматиды расходятся, попадая в разные дочерние клетки.
хромосомы человека
Кариоти́п - совокупность признаков (число, размеры, форма и т.д.) полного набора хромосом, присущий клеткам данного биологического вида (видовой кариотип ), данного организма (индивидуальный кариотип ) или линии (клона) клеток.
Для процедуры определения кариотипа могут быть использованы любые популяции делящихся клеток, для определения человеческого кариотипа используется либо одноядерные лейкоциты, извлечённые из пробы крови, деление которых провоцируется добавлением митогенов, либо культуры клеток, интенсивно делящихся в норме (фибробласты кожи, клетки костного мозга).
кариотип – диплойдный набор хромосом, свойтвенный соматическим клеткам организмов данного вида, являющийся видоспецифическим признаком и характеризующийся определённым числом и строением хромосом.
Хромосомный набор большинства клеток - диплоидный (2п) - это значит, что каждая хромосома имеет пару, т.е. гомологичную хромосому. Обычно диплоидный (2п) набор хромосом образуется в момент оплодотворения (одна из пары хромосом от отца, другая от матери). Некоторые клетки триплоидны (Зп), например клетки эндосперма.
Изменение числа хромосом в кариотипе человека может привести к различным заболеваниям. Наиболее частым хромосомным заболеванием у человека является синдром Дауна , обусловленный трисомией (к паре нормальных хромосом прибавляется еще одна такая же, лишняя) по 21-й хромосоме. Встречается этот синдром с частотой 1-2 на 1000.
Известны трисомии по 13-й хромосоме - Синдром Патау
, а также по 18-й хромосоме - синдром Эдвардса
, при которых жизнеспособность новорожденных резко снижена. Они гибнут в первые месяцы жизни из-за множественных пороков развития.
Достаточно часто у человека встречается изменение числа половых хромосом. Среди них известна моносомия Х (из пары хромосом присутствует только одна (Х0)) - это синдром Шерешевского-Тернера
. Реже встречается трисомия Х и синдром Клайнфельтера
(ХХУ, ХХХУ, ХУУ и т.д.)
6. Гиалоплазма. Органеллы, их классификация. Биологические мембраны.
гиалоплазма - часть цитоплазмы животных и растительных клеток, не содержащая структур, различимых в световом микроскопе.
Гиалоплазма (hyaloplasma; от греч. hyalinos - прозрачный) составляет примерно 53-55 % от общего объема цитоплазмы (cytoplasma), образуя гомогенную массу сложного состава. В гиалоплазме присутствуют белки, полисахариды, нуклеиновые кислоты, ферменты. При участии рибосом в гиалоплазме синтезируются белки, происходят различные реакции промежуточного обмена. В гиалоплазме располагаются также органеллы, включения и клеточное ядро.
Основная роль гиалоплазмы – объединение всех клеточных структур в отношении их химического взаимодействия и обеспечения транспортных биохимических процессов.
Органеллы (organellae) являются обязательными микроструктурами для всех клеток, выполняющими определенные жизненно важные функции. Различают мембранные и немембранные органеллы .
К мембранным органеллам , отграниченным от окружающей их гиалоплазмы мембранами, относятся эндоплазматическая сеть, комплекс Гольджи, лизосомы, пероксисомы, митохондрии.
Эндоплазматическая сеть представляет собой единую непрерывную структуру, образованную системой цистерн, трубочек и уплощенных мешочков. На электронных микрофотографиях различают зернистую (шероховатую, гранулярную) и незернистую (гладкую, агранулярную) эндоплазматическую сеть. Внешняя сторона зернистой сети покрыта рибосомами, незернистая лишена рибосом. Зернистая эндоплазматическая сеть синтезирует (на рибосомах) и транспортирует белки. Незернистая сеть синтезирует липиды и углеводы и участвует в их обмене (например, стероидные гормоны в корковом веществе надпочечников и клетках Лейдига (сустеноцитах) яичек; гликоген - в клетках печени). Одной из важнейших функций эндоплазматической сети является синтез мембранных белков и липидов для всех клеточных органелл.
комплекс Гольджи представляет собой совокупность мешочков, пузырьков, цистерн, трубочек, пластинок, ограниченных биологической мембраной. Элементы комплекса Гольджи соединены между собой узкими каналами. В структурах комплекса Гольджи происходят синтез и накопление полисахаридов, белково-углеводных комплексов, которые выводятся из клеток. Так образуются секреторные гранулы. Комплекс Гольджи имеется во всех клетках человека, кроме эритроцитов и роговых чешуек эпидермиса. В большинстве клеток комплекс Гольджи расположен вокруг или вблизи ядра, в экзокринных клетках - над ядром, в апикальной части клетки. Внутренняя выпуклая поверхность структур комплекса Гольджи обращена в сторону эндоплазматической сети, а внешняя, вогнутая, - к цитоплазме.
Мембраны комплекса Гольджи образованы зернистой эндоплазматической сетью и переносятся транспортными пузырьками. От внешней стороны комплекса Гольджи постоянно отпочковываются секреторные пузырьки, а мембраны его цистерн постоянно обновляются. Секреторные пузырьки поставляют мембранный материал для клеточной мембраны и гликокаликса. Таким образом обеспечивается обновление плазматической мембраны.
Лизосомы представляют собой пузырьки диаметром 0,2-0,5 мкм, содержащие около 50 видов различных гидролитических ферментов (протеазы, липазы, фосфолипазы, нуклеазы, гликозидазы, фосфатазы). Лизосомальные ферменты синтезируются на рибосомах зернистой эндоплазматической сети, откуда переносятся транспортными пузырьками в комплекс Гольджи. От пузырьков комплекса Гольджи отпочковываются первичные лизосомы. В лизосомах поддерживается кислая среда, ее рН колеблется от 3,5 до 5,0. Мембраны лизосом устойчивы к заключенным в них ферментам и предохраняют цитоплазму от их действия. Нарушение проницаемости лизосомальной мембраны приводит к активации ферментов и тяжелым повреждениям клетки вплоть до ее гибели.
Во вторичных (зрелых) лизосомах (фаголизосомах) происходит переваривание биополимеров до мономеров. Последние транспортируются через лизосомальную мембрану в гиалоплазму клетки. Непереваренные вещества остаются в лизосоме, в результате чего лизосома превращается в так называемое остаточное тельце высокой электронной плотности.
Митохондрии (mitochondrii), являющиеся «энергетическими станциями клетки», участвуют в процессах клеточного дыхания и преобразования энергии в формы, доступные для использования клеткой. Их основные функции - окисление органических веществ и синтез аденозинтрифосфорной кислоты (АТФ). Много крупных митохондрий в кардиомиоцитах, мышечных волокнах диафрагмы. Они расположены группами между миофибриллами, окружены гранулами гликогена и элементами незернистой эндоплазматической сети. Митохондрии являются органеллами с двойными мембранами (толщина каждой около 7 нм). Между наружной и внутренней митохондриальными мембранами расположено межмембранное пространство шириной 10-20 нм.
К немембранным органоидам относятся клеточный центр эукариотических клеток и рибосомы, имеющиеся в цитоплазме как эу- , так и прокариотических клеток.
Рибосома - это округлая рибонуклеопротеиновая частица диаметром 20-30 нм. Она состоит из малой и большой субъединиц, объединение которых происходит в присутствии матричной (информационной) РНК (мРНК). Одна молекула мРНК обычно объединяет несколько рибосом наподобие нитки бус. Такую структуру называют полисомой. Полисомы свободно располагаются в основном веществе цитоплазмы или прикреплены к мембранам шероховатой цитоплазматической сети. В обоих случаях они служат местом активного синтеза белка.
70S - рибосомы обнаруживаются у прокариот и в хлоропластах и митохондриях эукариот. 8OS-рибосомы, несколько более крупные, находятся в цитоплазме эукариот. В процессе синтеза белка рибосомы движутся вдоль мРНК. Процесс идет более эффективно, если вдоль мРНК движется не одна, а несколько рибосом. Такие цепи рибосом на мРНК называют полирибосомами, или полисомами.
МЕМБРАНЫ:
все мембраны образуют липопротеидные плёнки; имеют двойной слой липидов.
В составе мембран до 20% воды. липиды.
Мембраны состоят из липидов трех классов: фосфолипиды, гликолипиды и холестерол. Фосфолипиды и гликолипиды состоят из двух длинных гидрофобных углеводородных «хвостов», которые связаны с заряженной гидрофильной «головой». Холестерол придает мембране жесткость, занимая свободное пространство между гидрофобными хвостами липидов и не позволяя им изгибаться. Поэтому мембраны с малым содержанием холестерола более гибкие, а с большим - более жесткие и хрупкие.
Клеточные мембраны часто асимметричны, то есть слои отличаются по составу липидов, переход отдельной молекулы из одного слоя в другой (так называемый флип-флоп ) затруднен. Различается состав и ориентация мембранных белков.
Одна из важнейших функций биомембраны - барьерная. Например, мембрана пероксисом защищает цитоплазму от опасных для клетки пероксидов.
Еще одно важное свойство биомембраны - избирательная проницаемость.
Ядерные структуры
Простейшие синтаксические модели, являющиеся основой речевой деятельности, поскольку они используются для разнообразных трансформаций по требованиям контекста.
Словарь-справочник лингвистических терминов. Изд. 2-е. - М.: Просвещение . Розенталь Д. Э., Теленкова М. А. . 1976 .
Смотреть что такое "ядерные структуры" в других словарях:
ядерные структуры - простейшие синтаксические модели данного языка, являющиеся основой речевой деятельности в том смысле, что пользующиеся данным языком подвергают эти модели разнообразным трансформациям в зависимости от ее требований контекста. Ср. ядерные… …
Превращения ат. ядер при вз ствии с ч цами, в т. ч. с g квантами или друг с другом. Для осуществления Я. р. необходимо сближение ч ц (двух ядер, ядра и нуклона и т. д.) на расстояние 10 13 см. Энергия налетающих положительно заряж. ч ц должна… … Физическая энциклопедия
ядерные фибриллы - Нитевидные внутриядерные структуры, являющиеся фрагментами ядерного скелета [Арефьев В.А., Лисовенко Л.А. Англо русский толковый словарь генетических терминов 1995 407с.] Тематики генетика EN nuclear fibrils … Справочник технического переводчика
Превращения атомных ядер при взаимодействии с элементарными частицами, γ квантами или друг с другом. Для осуществления Я. р. необходимо сближение частиц (двух ядер, ядра и нуклона и т. д.) на расстояние Ядерные реакции 10 13 см. Энергия… …
Обмен веществами между ядром и цитоплазмой клетки осуществляется посредством ядерных пор транспортных каналов, пронизывающих двухслойную ядерную оболочку. Переход молекул из ядра в цитоплазму и в обратном направлении называется ядерно… … Википедия
Сильное взаимодействие (цветовое взаимодействие, ядерное взаимодействие) одно из четырёх фундаментальных взаимодействий в физике. Сильное взаимодействие действует в масштабах атомных ядер и меньше, отвечая за притяжение между нуклонами в ядрах и … Википедия
Nuclear fibrils ядерные фибриллы. Hитевидные внутриядерные структуры, являющиеся фрагментами ядерного скелета
ядерные предложения - простейшие синтаксические построения данного языка, в которых предметы обозначены существительными, процессы глаголами, а признаки прилагательными и наречиями, от которых путем серии трансформаций образуются поверхностные структуры … Толковый переводоведческий словарь
ядерные реакции - превращение атомов ядер при соударении с другими ядрами, элементарными частицами или гамма квантами. При бомбардировке тяжелых ядер более легкими получены все трансурановые элементы. Сокращенно ядерную реакцию, например, типа… … Энциклопедический словарь по металлургии
Ядерные процессы, в которых вносимая в атомное ядро энергия передаётся преимущественно одному или небольшой группе нуклонов (См. Нуклоны). П. я. р. многообразны, они вызываются всевозможными налетающими частицами (от γ квантов до… … Большая советская энциклопедия
Книги
- Инновационная деятельность в атомной отрасли (на примере стратегии развития ядерных топливных циклов, включая инновационные). Книга 1. Основные принципы инновационной политики , А. В. Путилов, А. Г. Воробьев, М. Н. Стриханов. В учебном пособии раскрываются роль и место инноваций в общественном развитии на примере атомной отрасли; цели и задачи национальной инновационной политики. Рассмотрены инструменты…
- Введение в физику микромира. Физика частиц и ядер , Л. И. Сарычева. В настоящей книге представлены основные характеристики фундаментальных и элементарных частиц и процессы, происходящие с ними в различных типах взаимодействий. Описана современная…
Ядро представляет собой обязательную часть клетки у многих одноклеточных и всех многоклеточных организмов.
Рис. 1.
Оно содержит ядерные гены, и соответственно выполняет 2 главные функции:
1. Хранение и воспроизведение генетической информации;
2. Регуляция процессов обмена веществ, протекающих в клетке.
По наличию или отсутствию в клетках оформленного ядра все организмы делятся на прокариотические и эукариотические. Основное отличие заключается в степени обособления генетического материала (ДНК) от цитоплазмы и в образовании у эукариот сложных ДНК-содержащих структур-хромосом. Клетки эукариот содержат оформленные ядра. Клетки прокариот не имеют морфологически оформленного ядра.
Путем реализации заключенной в генах наследственной информации ядро управляет белковыми синтезами, физиологическими и морфологическими процессами в клетке. Функции ядра осуществляются в тесном взаимодействии с цитоплазмой.
Впервые ядро наблюдал Я. Пуркине (1825) в яйцеклетке курицы. Ядра растительных клеток были описаны Р. Броуном (1831-33), который наблюдал в них шарообразные структуры. Ядра животных клеток были описано Т. Шванном (1838-39 гг.)
Размеры ядра колеблются от 1 мкм (у некоторых простейших) до 1 мм (в яйцах некоторых рыб и земноводных). В состав большинства эукариотических клеток входит одно ядро. Однако, встречаются и многоядерные клетки (поперечнополосатые мышечные волокна и т.д.). В состав клеток инфузории, например, входит 2 ядра (макронуклеус и микронуклеус). Встречаются и полиплоидные клетки, в которых произошло увеличение наборов хромосом.
Форма ядра может быть различной (сферической, эллипсовидной, неправильной и т.д.) и зависит от формы клетки.
Между объемом ядра и объемом цитоплазмы существует взаимосвязь. Более молодые клетки обычно имеют более крупные ядра. Положение ядра в клетке может меняться по мере дифференцировки или накопления питательных веществ.
Ядро окружено ядерной мембраной, которая является двухслойной и содержит ядерные поры, расположенные на равном расстоянии друг от друга.
В состав интерфазного ядра входят кариоплазма, хроматин, ядрышки, а также синтезируемые в ядре структуры (перихроматиновые фибриллы, перихроматиноые гранулы, интерхроматиновые гранулы). Во время активных фаз деления ядра происходит спирализация хроматина и образование хромосом.
Структура ядра неоднородна. Имеются более спирализованные гетерохроматиновые участки (ложные или хроматиновые ядрышки). Остальные участки - эухроматиновые. Удельный вес ядра выше, чем у остальной цитоплазмы. Среди ядерных структур наибольшим весом обладает ядрышко. Вязкость ядра выше, чем вязкость цитоплазмы. Если ядерная оболочка разрывается и кариоплазма выходит наружу, происходит спадание ядра без всяких признаков реконструкции.

Рис. 2.

Рис. 3.
Ядерная оболочка состоит из двух мембран, причем наружная является продолжением мембраны эндоплазматического ретикулума. Липидный бислой внутренней и наружной ядерных мембран соединяются в ядерных порах. Две сети нитевидных промежуточных фибрилл (цветные линии) обеспечивают механическую прочность ядерной оболочки.Фибриллы внутри ядра образуют подстилающую ядерную ламину (по Альбертсу).
Ядерная оболочка непосредственно связана с эндоплазматическим ретикулумом. С обеих сторон к ней прилегают сетеподобные структуры, состоящие из промежуточных филаментов. Сетеподобная структура, которая выстилает внутреннюю ядерную мембрану называется ядерной ламиной.

Рис. 4.
Ядерная оболочка
Эта структура характерна для всех эукариотических клеток. Ядерная оболочка состоит из внешней и внутренней липопротеидных мембран, толщина которых составляет 7-8 нм. Липопротеидные мембраны разделены перинуклеарным пространством шириной от 20 до 60 нм. Ядерная оболочка ограничивает ядро от цитоплазмы.
Ядерная оболочка пронизана порами, диаметр которых составляет 60-100 нм. По краю каждой поры находится плотное вещество (аннулус). По границе округлого отверстия в ядерной оболочке располагаются три ряда гранул, по 8 штук в каждом: один ряд лежит со стороны ядра, другой - со стороны цитоплазмы, третий расположен в центральной части пор. Размер гранул около 25 нм. От этих гранул отходят фибриллярные отростки, в просвете поры имеется центральный элемент диаметром 15-20 нм, соединенный с аннулусом радиальными фибриллами. Вместе эти структуры образуют поровый комплекс, который регулирует прохождение макромолекул через поры.
Внешняя ядерная мембрана может переходить в мембраны эндоплазматической сети. На внешней ядерной мембране обычно располагается большое количество рибосом. У большинства животных и растительных клеток внешняя мембрана ядерной оболочки не представляет собой идеально ровную поверхность - она может образовывать различной величины выпячивания или выросты в сторону цитоплазмы.
Число ядерных пор зависит от метаболической активности клеток: чем выше синтетические процессы в клетках, тем больше пор на единицу поверхности клеточного ядра.
С химической точки зрения, в состав ядерной оболочки входит ДНК (0-8%), РНК (3-9%), липиды (13-35%) и белки (50-75%).
Что касается липидного состава ядерной мембраны, то он сходен с химическим составом мембран ЭПС (эндоплазматической сети). В ядерных мембранах наблюдается низкое содержание холестерина и высокое содержание фосфолипидов.
Белковый состав мембранных фракций очень сложен. Среди белков обнаружен ряд ферментов, общих с ЭР (например, глюкозо-6-фосфатаза, Mg-зависимая АТФаза, глютамат-дегидрогеназа и др.) не обнаружена РНК-полимераза. Тут выявлены активности многих окислительных ферментов (цитохромоксидазы, НАДН-цитохром-с-редуктазы) и различных цитохромов.
Среди белковых фракций ядерных мембран встречаются основные белки типа гистонов, что объясняется связью участков хроматина с ядерной оболочкой.
Ядерная оболочка проницаема для ионов, веществ с малым молекулярным весом (сахара, аминокислоты, нуклеотиды). Из ядра в цитоплазму происходит транспорт РНК.
Ядерная оболочка является барьером, ограничивающим содержимое ядра от цитоплазмы и препятствующим свободному доступу в ядро крупных биополимеров.

Рис. 5. Ядерная оболочка отделяет ядро от цитоплазматических органелл. На этой электронной микрофотографии представлен тонкий срез ооцита морского ежа, ядро которого окрашивается необычайно равномерно, а цитоплазма плотно забита органеллами. (По Альбертсу)
Кариоплазма
Кариоплазма или ядерный сок - это содержимое клеточного ядра, в которое погружены хроматин, ядрышки, внутриядерные гранулы. После экстракции хроматина химическими агентами в кариоплазме сохраняется так называемый ядерный матрикс. Этот комплекс не представляет собой какую-то чистую фракцию, сюда входят компоненты и ядерной оболочки, и ядрышка, и кариоплазмы. С ядерным матриксом оказались связаны как гетерогенная РНК, так и часть ДНК. Матрикс ядра играет важную роль не только в поддержании общей структуры интерфазного ядра, но и может участвовать в регуляции синтеза нуклеиновых кислот.
Хроматин
Клеточное ядро является вместилищем почти всей генетической информации клетки, поэтому основное содержимое клеточного ядра -- это хроматин: комплекс дезоксирибонуклеиновой кислоты (ДНК) и различных белков. В ядре и, особенно, в митотических хромосомах, ДНК хроматина многократно свернута, упакована особым образом для достижения высокой степени компактизации.
Ведь все длинные нити ДНК необходимо уложить в клеточное ядро, диаметр которого всего несколько микрометров. Эта задача решается последовательной упаковкой ДНК в хроматине с помощью специальных белков. Основная масса белков хроматина -- это белки гистоны, входящие в состав глобулярных субъединиц хроматина, называемых нуклеосомами. Хроматин представляет собой нуклеопротеидные нити, входящие в состав хромосом. Термин «хроматин» был введен В.Флеммингом (1880). Хроматин - это дисперсное состояние хромосом в интерфазе клеточного цикла. Основными структурными компонентами хроматина являются: ДНК (30-45%), гистоны (30-50%), негистоновые белки (4-33%). Существует 5 типов белков-гистонов, входящих в состав хроматина (Н1, Н2А, Н2В, Н3 и Н4). Белок Н1 слабо связан с хроматином.
По своей морфологии хроматин напоминает структуру «бус», состоящих из нуклеосом (частиц диаметром около 10 нм). Нуклеосома- это сегмент ДНК длиной 200 пар оснований, навитый на белковую сердцевину, которая состоит из 8 молекул белков-гистонов (Н2А, Н2В, Н3 и Н4). Каждая нуклеосома маскирует 146 пар оснований. Нуклеосома представляет собой цилиндрическую частицу, состоящую из 8 молекул гистонов, диаметром около 10 нм, на которую «намотано» чуть менее двух витков нити молекулы ДНК. Все белки-гистоны, кроме Н1, входя в состав сердцевины нуклеосомы. Белок Н1 вместе с ДНК связывает отдельные нуклеосомы между собой (этот участок называется линкерная ДНК). В электронном микроскопе такой искусственно деконденсированный хроматин выглядит как «бусины на нитке». В живом ядре клетки нуклеосомы плотно объединены между собой с помощью еще одного линкерного гистонового белка, образуя так называемую элементарную хроматиновую фибриллу, диаметром 30 нм. Другие белки, негистоновой природы, входящие в состав хроматина обеспечивают дальнейшую компактизацию, т. е. укладку, фибрилл хроматина, которая достигает своих максимальнах значений при делении клетки в митотических или мейотических хромосомах. В ядре клетки хроматин присутствует как в виде плотного конденсированного хроматина, в котором 30 нм элементарные фибриллы упакованы плотно, так и в виде гомогенного диффузного хроматина. Количественное соотношение этих двух видов хроматина зависит от характера метаболической активности клетки, степени ее дифференцированности. Так, например, ядра эритроцитов птиц, в которых не происходит активных процессов репликации и транскрипции, содержат практически только плотный конденсированный хроматин. Некоторая часть хроматина сохраняет свое компактное, конденсированное состояние в течение всего клеточного цикла -- такой хроматин называется гетерохроматином и отличается от эухроматина рядом свойств.
Спирализованные участки хромосом инертны в генетическом отношении. Передачу генетической информации осуществляют деспирализованные участки хромосом, которые в силу своей малой толщины не видны в световой микроскоп. В делящихся клетках все хромосомы сильно спирализуются, укорачиваются и приобретают компактные размеры и форму.
Хроматин интерфазных ядер представляет собой несущие ДНК тельца (хромосомы), которые теряют в это время свою компактную форму, разрыхляются, деконденсируются. Степень такой деконденсации хромосом может быть различной в ядрах разных клеток. Когда хромосома или ее участок полностью деконденсирован, тогда эти зоны называют диффузным хроматином. При неполном разрыхлении хромосом в интерфазном ядре видны участки конденсированного хроматина (иногда называемого гетерохроматин). Показано, что степень деконденсации хромосомного материала в интерфазе может отражать функциональную нагрузку этой структуры. Чем более диффузен хроматин интерфазного ядра, тем выше в нем синтетические процессы. Падение синтеза РНК в клетках обычно сопровождается увеличением зон конденсированного хроматина.
Максимально конденсирован хроматин во время митотического деления клеток, когда он обнаруживается в виде плотных телец - хромосом. В этот период хромосомы не несут никаких синтетических нагрузок, в них не происходит включение предшественников ДНК и РНК.

Рис. 6.
Нуклеосомные частицы состоят из двух полных витков ДНК (83 нуклеотидных пары на виток), закрученных вокруг кора, представляющего собой гистоновый октамер, и соединяются между собой линкерной ДНК. Нуклеосом-ная частица выделена из хроматина путем ограниченного гидролиза линкерных участков ДНК микрококковой нуклеазой. В каждой нуклеосомнои частице фрагмент двойной спирали ДНК, имеющий в длину 146 пар оснований, закручен вокруг гистонового кора. Этот белковый кор содержит по две молекулы каждого из гистонов Н2А, Н2В, НЗ и Н4. Полипептидные цепи гистонов насчитывают от 102 до 135 аминокислотных остатков, а общий вес октамера составляет приблизительно 100000 Да. В деконденсированной форме хроматина каждая «бусина» связана с соседней частицей нитевидным участком линкерной ДНК (по Альбертсу).

Рис. 7.

Рис. 8.
Показаны три нити хроматина, на одной из которых две молекулы РНК-полимеразы транскрибируют ДНК. Большая часть хроматина в ядре высших эукариот не содержит активных генов, и, следовательно, свободна от РНК-транскриптов. Следует отметить, что нуклеосомы имеются как в транскрибируемых, так и в нетранскрибируемых областях, и что они связаны с ДНК непосредственно перед и сразу же за движущимися молекулами РНК-полимераз. (по Альбертсу) .

Рис. 9.
А. Вид сверху. Б. Вид сбоку.
При таком типе упаковки на нуклеосому приходится одна молекула гистона Н1 (не указано). Хотя место прикрепления гистона Н1 к нуклеосоме определено, расположение молекул Н1 на этой фибрилле неизвестно (по Альбертсу).
Белки хроматина
Гистоны - сильноосновные белки. Их щелочность связана с их обогащенностью основными аминокислотами (главным образом лизином и аргинином). Эти белки не содержат триптофана. Препарат суммарных гистонов можно разделить на 5 фракций:
Н 1 (от английского histone) - богатый лизином гистон, мол. Масса 2100;
Н 2а - умеренно богатый лизином гистон, масса 13 700;
Н 2б - умеренно богатый лизином гистон, масса 14 500;
Н 4 - богатый аргинином гистон, масса 11 300;
Н 3 - богатый аргинином гистон, масса 15 300.
В препаратах хроматина эти фракции гистонов обнаруживаются в приблизительно равных количествах, кроме Н 1 , которого примерно в 2 раза меньше любой из других фракций.
Для молекул гистонов характерно неравномерное распределение основных аминокислот в цепи: обогащенные положительно заряженными аминогруппами наблюдается на концах белковых цепей. Эти участки гистонов связываются с фосфатными группировками на ДНК, в то время как сравнительно менее заряженные центральные участки молекул обеспечивают их взаимодействие между собой. Таким образом, взаимодействие между гистонами и ДНК, приводящее к образованию дезоксирибонуклеопротеинового комплекса, носит ионный характер.
Гистоны синтезируются на полисомах в цитоплазме, этот синтез начинается несколько раньше редупликации ДНК. Синтезированные гистоны мигрируют из цитоплазмы в ядро, где и связываются с участками ДНК.
Функциональная роль гистонов не вполне ясна. Одно время считалось, что гистоны являются специфическими регуляторами активности ДНК хроматина, но одинаковость строения основной массы гистонов говорит о малой вероятности этого. Более очевидна структурная роль гистонов, которая обеспечивает не только специфическую укладку хромосомной ДНК, но и играет роль в регуляции транскрипции.

Рис. 10.
Нуклеосомные частицы состоят из двух полных витков ДНК (83 нуклеотидных пары на виток), закрученных вокруг кора, представляющего собой гистоновый октамер, и соединяются между собой линкерной ДНК. Нуклеосом-ная частица выделена из хроматина путем ограниченного гидролиза линкерных участков ДНК микрококковой нуклеазой. В каждой нуклеосомнои частице фрагмент двойной спирали ДНК, имеющий в длину 146 пар оснований, закручен вокруг гистонового кора. Этот белковый кор содержит по две молекулы каждого из гистонов Н2А, Н2В, НЗ и Н4. Полипептидные цепи гистонов насчитывают от 102 до 135 аминокислотных остатков, а общий вес октамера составляет приблизительно 100000 Да. В деконденсированной форме хроматина каждая «бусина» связана с соседней частицей нитевидным участком линкерной ДНК.
Негистоновые белки - наиболее плохо охарактеризованная фракция хроматина. Кроме ферментов, непосредственно связанных с хроматином (ферменты, ответственные за репарацию, редубликацию, транскрипцию и модификации ДНК, ферменты модификации гистонов и других белков), в эту фракцию входит множество других белков. Весьма вероятно, что часть негистоновых белков представляет собой специфические белки - регуляторы, узнающие определенные нуклеотидные последовательности в ДНК.
РНК хроматина составляет от 0,2 до 0,5% от содержания ДНК. Эта РНК представляет собой все известные клеточные типы РНК, находящиеся в процессе синтеза или созревания в связи с ДНК хроматина.
В составе хроматина могут быть обнаружены липиды до 1 % от весового содержания ДНК, их роль в структуре и функционировании хромосом остается неясной.
В химическом отношении препараты хроматина представляют собой сложные комплексы дезоксирибонуклеопротеидов, в состав которых входит ДНК и специальные хромосомные белки - гистоны. В составе хроматина обнаружено также РНК. В количественном отношении ДНК, белок и РНК находятся как 1:1,3:0,2. О значении РНК в составе хроматина еще нет достаточно однозначных данных. Возможно, что эта РНК представляет собой сопутствующую препарату функцию синтезирующейся РНК и поэтому частично связанной с ДНК или это особый вид РНК, характерный для структуры хроматина.
ДНК хроматина
В препарате хроматина на долю ДНК приходится обычно 30-40%. Эта ДНК представляет собой двухцепочечную спиральную молекулу. ДНК хроматина обладает молекулярной массой 7-9*10 6 . Такую сравнительно малую массу ДНК из препаратов можно объяснить механическими повреждениями ДНК в процессе выделения хроматина.
Общее количество ДНК, входящее в ядерные структуры клеток, в геном организмов, колеблется от вида к виду. Сравнивая количество ДНК на клетку у эукариотических организмов, трудно уловить какие-либо корреляции между степенью сложности организма и количеством ДНК на ядро. Примерно одинаковое количество ДНК имеют различные организмы, как лен, морской еж, окунь (1,4-1,9 пг) или рыба голец и бык (6,4 и 7 пг).
У некоторых амфибий в ядрах количество ДНК больше, чем в ядрах человека, в 10-30 раз, хотя генетическая конституция человека несравненно сложнее, чем у лягушек. Следовательно, можно предполагать, что избыточное количество ДНК у более низко организованных организмов либо не связано с выполнением генетической роли, либо число генов повторяется то или иное число раз.
Сателлитная ДНК, или фракция ДНК с часто повторяющимися последовательностями, может участвовать в узнавании гомологичных районов хромосом при мейозе. По другим предположениям, эти участки играют роль разделителей (спейсеров) между различными функциональными единицами хромосомной ДНК.
Как оказалось, фракция умеренно повторяющихся (от 10 2 до 10 5 раз) последовательностей принадлежит к пестрому классу участков ДНК, играющих важную роль в обменных процессах. В эту фракцию входят гены рибосомных ДНК, многократно повторенные участки для синтеза всех тРНК. Более того, некоторые структурные гены, ответственные за синтез определенных белков, также могут быть многократно повторены, представлены многими копиями (гены для белков хроматина - гистонов).
Ядрышко
Ядрышко (нуклеола) - это плотное тельце внутри ядра большинства клеток эукариот. Состоит из рибонуклеопротеидов - предшественников рибосом. Обычно в клетке одно ядрышко, реже много. В ядрышке выделяют зону внутриядрышкового хроматина, зону фибрилл и зону гранул. Ядрышко-это не постоянная структура в клетках эукариот. При активном митозе ядрышки распадаются, а затем синтезируются вновь. Основная функция ядрышек-синтез РНК и субъединиц рибосом.
В ядрышке выделяют зону внутриядрышкового хроматина, зону фибрилл и зону гранул. Ядрышко не является самостоятельным органоидом клетки, лишено мембраны и образуется вокруг участка хромосомы, в котором закодирована структура рРНК (ядрышковый организатор), на нем синтезируется рРНК; кроме накопления рРНК в ядрышке формируются рибосомы, которые затем перемещаются в цитоплазму. Т.о. ядрышко - это скопление рРНК и рибосом на разных этапах формирования.
Основной функцией ядрышка является синтез рибосом (в этом процессе принимает участие РНК-полимераза I)








